--David Duncan and Jack Widholm
For studies of somaclonal variation using type I regenerable maize callus, it is useful to obtain from the callus as many regenerated plants as possible. This goal is desirable primarily because not all regenerated plants survive or set seed and a large number of plants are needed to have enough plant material to conduct a valid experiment. Not all regenerated plantlets, however, develop at the same rate. Consequently, regenerated plantlets may be harvested from a given Petri dish of regeneration medium over about a 30d period. Poor plant survival and a lack of synchrony to plant regeneration has prompted studies aimed at increasing the efficiency of plant regeneration from maize callus cultures.
Callus from the self-pollinated inbred Mo17 was initiated and maintained as described by Duncan, Williams, Zehr and Widholm (Planta 165:322). After maintenance for 205d, plants were regenerated from the callus as described by Duncan and Widholm (Plant Cell Reports 7:452). One half of the callus was placed in a 12h light/dark cycle and the other half was placed in continuous light. Plants were removed from regeneration medium at three 10-d intervals.
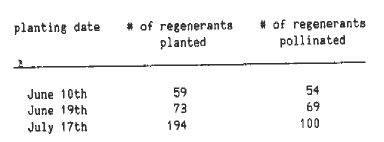
Six hundred five shoots were removed from regeneration medium and placed in individual culture tubes and from these, 326 plants were placed in the greenhouse. Many of the shoots in tubes that were not placed in the greenhouse developed so late and so slowly that it was futile to attempt to move them to the field. Plants were taken to the field after approximately two weeks in the greenhouse. Planting dates were June 10th, June 19th, and July 17th (1987).
Table 1. The number of regenerated plants moved to the field and pollinated for each planting date.
Of the 336 plants taken to the greenhouse, 326 were transplanted to the field and 223 of these plants were pollinated (Table 1). Of the 103 plants not pollinated, 12.6% were from the first and second planting and 87.4% were from the third planting. More than 50% of plants from the third planting were pollinated and viable seed was obtained and used the following year. However, many of the plants in the third planting matured too late in the season to be pollinated. Whether this late maturity was due to somaclonal variation (such as has been previously documented; Zehr, Williams, Duncan and Widholm, 1987) or environmental stress could not be determined.
Forty-two plants were found with tassel ears and all of these were from the third planting. No tassel ears were found on control plants derived from seed that had been germinated in the greenhouse and transplanted to the field at the same time.
Three plants from the second planting and five plants from the third planting produced R1 seed with a shrunken phenotype but this was not found in seed from plants of the first planting. Germination of R1 seed decreased with planting date (third planting, 60%; second planting, 69%; first planting, 75%). This lowered germination may be the result of a shorter grain fill period for the later plantings.
The 12h light treatment delayed plant development by about 10d, which, because of the limited length of the growing season, resulted in fewer plants being moved to the field and fewer pollinations of these plants being made (Table 2).
Table 2. The effect of light on plant regeneration from regenerable maize callus.
For instance, no plants from the 12h treatment were included in the first planting. Also, of the 42 plants with tassel ears 71.0% were from the 12h light treatment. These results suggest that the 12h light/dark cycle may be detrimental to effective regeneration of normal plants from maize callus cultures.
There are obvious environmental effects resulting from the different planting dates, for instance, small plants and poor grain fill seen in material planted later. These effects may also carry over to the progeny, for instance, poor germination and seedling vigor. However, the regenerated plants that were planted later also appeared more abnormal, as indicated by the prevalence of tassel ears in the last planting.
The last planted regenerants were so planted because they either differentiated
later or slower, or they grew slower. To date we do not know if this variation
in shoot development resulted from segregation in time of somaclonal variants
(i.e., the most normal plants developing the fastest) or from changes with
time in culture condition that adversely affect normal shoot development.
We can conclude, however, that harvesting all of the regenerated shoots
from a Petri dish of regeneration medium may not be the most efficient
means of plant regeneration. Instead, using a large quantity of callus
and only the first few regenerated plants found on regeneration medium
may be the most efficient method of producing the most normal plant material.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}