Howard Hughes Medical Institute
University of Utah
--Christiane Fauron and Marie Havlik
The mitochondrial genome of different lines varies in size and organization. The physical map of the normal cytoplasm in a Wf9 or B37 nuclear background (Lonsdale et al., Nucl. Acid. Res. 16:9249, 1984, Fauron and Havlik, Nucl. Acid. Res. 16:10395, 1988) reveals a sequence complexity of 570kb. However, the physical map of the cytoplasmic male sterile type Texas (cms-T) in a Wf9 and B37 nuclear background reveals a sequence complexity of 540kb (Fauron et al., Mol. Gen. Genet., in press, 1989a, Fauron unpublished). For both genomes the entire sequence complexity can be represented on a master chromosome or alternatively as a multipartite structure via recombination at repeated sequences.
Using the three restriction enzymes BamHI, XhoI, and SmaI, 350 and 339 sites have been located in the N and cms-T map respectively. A detailed comparison of N and cms-T mitochondrial genomes by hybridization studies (genotype B37, Fauron and Havlik, Curr. Genet., in press, 1989b) led to the following observations:
1. Because the repeated sequences are mostly different between N and cms-T, recombination generates a rather different population of submolar circular molecules.
2. A complex pattern of sequence permutations (involving sequences as small as 1kb, as large as 95kb) between the two genomes indicates that at least thirty events are needed to explain the sequence rearrangement.
3. The sequences not shared between the two genomes (70kb in N and 40kb in cms-T) can be accounted for by the presence of different repeated sequences, the absence of the integrated form of plasmids R1 and R2 in cms-T, the presence and absence of chloroplast sequences and a number of unidentified sequences that are specific to N or to cms-T.
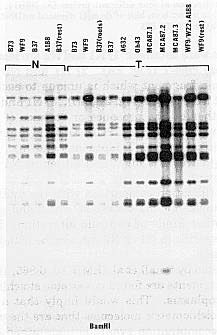
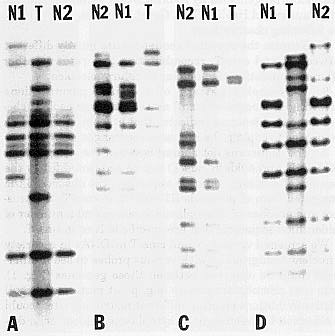
We screened various N and cms-T mtDNAs in a variety of nuclear backgrounds with various probes to see if differences could be observed between those genomes (Fig. 1). some rare microheterogeneity (e.g. point mutations, small deletions/additions creating different restriction sites) could be observed between some N cytoplasms. However, a distinct second class of normal mtDNA organization could also be identified (genotype A188, Ky21, W182B) when most of the cosmid clones used as probes give an hybridization pattern rather different from the known N or cms-T mtDNA, suggesting that the physical map is different as is the sequence complexity. This second class of normal maize mitochondrial genome is called N2 versus the N1 notation that is going to be used for the already published N map (Wf9, B37). An example of the hybridization studies is given in Figure 2. Depending on the clone used as a probe, four different results were obtained: 1) N1 and N2 are identical, but different from cms-T (Fig. 2A). 2) N1 and N2 differ by more than one fragment which is unique to each genome and different from cms-T (Fig. 2B, 2D). 3) N1 and N2 differ from each other by the presence or absence of fragment also shared with cms-T (Fig. 2C). 4) N2 is different from N1 but identical to cms-T (data not shown). Also, N2 contains sequences not identified either in N1 or cms-T (data not shown).
Figure
1. Hybridization of the cosmid clone Tu1E3 to a Southern blot containing
BamHI digested and mtDNA of N and cms-T in a variety of nuclear
backgrounds as indicated on the top of the picture. NA188 is different
from the other Ns. No consistent difference was observed between the T
cytoplasms.
[rest] indicates the cytoplasms containing the dominant
nuclear genes Rf1 and Rf2.
Figure 2. Hybridization of four different probes (A: N8B11, B: N7C9, C: N8A1, D: N5G8) to a Southern blot of BamHI digested mtDNA from NB37 (N1), NA188 (N2) and TB37 (T).
More hybridization comparison with the mtDNA from the other cytoplasmic male steriles, cms-S and cms-C, revealed yet other kinds of genomic alteration. However, some bands specific to N1 or N2 can also be found in either cms-C or cms-S.
As postulated by Small et al. (EMBO J. 6:865, 1987) some restriction
fragments are found in variable stoichiometry in different cytoplasms.
This would imply that a common pool of substoichiometric molecules that
are the product of infrequent recombination events is maintained at a very
low level in all of the maize cytoplasms. A mechanism of differential amplification
would determine which one of those substoichiometric molecules will be
amplified in the various cytoplasms.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}