University of Minnesota
RALEIGH, NORTH CAROLINA
North Carolina State University
its completely unimportant, that's why its so interesting
--John Doebley and Paul H. Sisco
During the course of a survey of variation in the chloroplast genome of maize and the teosintes, we examined the chloroplast genomes of the maize male sterile cytoplasms cms-C, S and T. The question in mind was whether (1) these cytoplasms were similar to the fertile cytoplasms of maize for their chloroplast genome or (2) they were foreign cytoplasms (probably teosinte cytoplasms) which have become incorporated into a maize nuclear background. If the former is the case, then it is probable that the male sterile trait arose within maize, most likely as a result of mutations in the mitochondrial genome (Model 1). If the latter is true, then male sterility may have originated because of incompatibility between the foreign cytoplasm and the maize nuclear genome (Model 2).
Chloroplast DNA or total cellular DNA preparations of maize and teosinte were restricted with 21 restriction enzymes, electrophoresed, blotted and probed with cloned portions of the chloroplast genome representing 87% of its length. The analyses included 86 accessions of maize, mostly U.S. and Latin American landraces, and 74 accessions of teosinte. Eight cpDNA types were defined by the presence/absence of restriction sites or insertion/deletion events. Four of these eight types were very distinct from maize (14 to 16 restriction site differences) and were restricted to the teosintes, Zea diploperennis, Z. perennis and Z. luxurians. The remaining five types were quite similar to one another (only 1 or 2 site differences) and were found in maize and the Mexican annual teosintes, Z. mays subsp. mexicana and subsp. parviglumis (Table 1). The chloroplast genome types of cms-C and T were the same as those found in the majority of the maize accessions (fertile cytoplasms) and subsp. parviglumis, the teosinte that cytological and isozymic data indicate was the progenitor of maize (Table 1, type E). This implies that the origin of the male sterility of these cytoplasms fits Model 1 (above). The chloroplast genome type of cms-S (Table 1, type A), however, was not found in any other type of maize but was present in three collections of Z. mays subsp. mexicana (Central Plateau Teosinte) from Copandiro, Michoacan, Mexico. This chloroplast genome type is distinguished from the common type of maize by its lack of both an 80bp deletion and a single EcoRI site typical of normal maize (type E). The 80bp deletion had been previously observed by Pring and Levings (Genetics 89:121-136, 1978). To confirm that this teosinte and cms-S actually contain the same cytoplasm, we isolated mitochondrial DNA from this teosinte and observed that it contains the S-1 and S-2 plasmids typical of cms-S. Restriction fragment banding patterns of mitochondrial DNA revealed that Copandiro teosinte belongs to cms-S subgroup CA as defined by Sisco et al. (Theor. Appl. Genet. 71:5-15, 1985). This is the most common of the cms-S subgroups. Thus, there seems to be little question that this teosinte and cms-S share the same cytoplasm.
Table 1. Numbers of accessions of maize and Mexican annual teosinte
possessing each of five distinct chloroplast DNA genotypes.
| Taxon |
|
||||
| A | B | C | D | E | |
| subsp. mexicana | 3 | 4 | 24 | - | - |
| subsp. parviglumis | - | 1 | 16 | 12 | 3 |
| subsp. mays | |||||
| landraces | - | 32 | 8 | - | 45 |
| cms-C | - | - | - | - | 1 |
| cms-S | 1 | - | - | - | - |
| cms-T | - | - | - | - | 1 |
a Using Zea diploperennis as an outgroup, cpDNA type A would be judged most primitive (the first to diverge from the others). Type B is the next derived form, distinguished by a deletion mutation (LM-3). Types C, D and E are the most advanced, each possessing LM-3 plus additional restriction site loss/gains. Type C has an unique EcoRI site; Type D has an unique CfoI site; Type E has another unique EcoRI site.
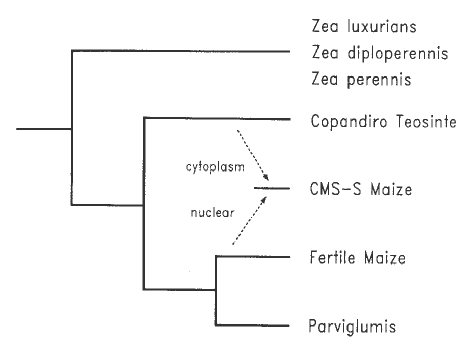
There are two possible explanations for the presence of the same cytoplasm in cms-S and Copandiro teosinte: (1) cms-S represents a domestication from Copandiro teosinte (subsp. mexicana), while the fertile cytoplasm types represent a separate domestication from subsp. parviglumis; or (2) introgression was involved. Of these alternatives, introgression seems more plausible as it is consistent with other data which suggest that maize was domesticated only once and that subsp. parviglumis (not subsp. mexicana) was the ancestral teosinte (see Doebley, Goodman & Stuber, Econ. Bot. 41:234-246, 1987). Accepting the introgression hypothesis, the next question is did the cms-S cytoplasm originate in maize and subsequently become transferred into Copandiro teosinte or the reverse. Again, if one accepts the evidence (1) that teosinte existed before maize, (2) that maize represents only a recent branchlet in the evolution of Zea, and (3) that subsp. parviglumis was the progenitor of maize (see Doebley, Goodman & Stuber, Econ. Bot. 41:234-246, 1987), then the direction of introgression must have been from Copandiro teosinte into maize because the original domesticated maize would have had the cytoplasm type of subsp. parviglumis. cms-S cytoplasm is also found in race Cónico Norteño maize of the Mexican Central Plateau (Weissinger et al., Genetics 104:365-379, 1983). This race might have been the link between Copandiro teosinte and the cms-S cytoplasms of the U.S. A scenario for the origin of cms-S cytoplasm is graphically summarized in Fig. 1.
Figure 1. A phylogeny showing the presumed origin of cms-S maize by the introgression of Copandiro teosinte cytoplasm (=cms-S cytoplasm) into the maize nuclear background.
This interpretation of the facts suggests that the origin of male sterility
in cms-S maize follows model 2 (above). If this is true, Copandiro teosinte
may possess restorer genes and may even have been the source of the restorer
genes for cms-S that are found in maize. A question we do not know the
answer to is whether Copandiro teosinte exhibits the male sterile trait,
although this seems unlikely as that would inhibit or even prevent its
survival as a wild plant.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}