--J. W. Higginbotham*, J. S. C. Smith and O. S. Smith
*Also affiliated with Cold Spring Harbor Lab.
Before two-dimensional corn seedling protein profiles are routinely used to describe inbred genotypes and to determine genetic associations between inbreds, the contribution of slight differences in the development of seedlings to the variation present in the protein profiles needs to be understood to insure adequate stability of the profile.
This experiment was designed to test the impact of slightly varying total imbibition periods on the protein profile relative to the impact of different genotypes. One lot each of Pioneer Hi-Bred proprietary inbred lines G39 and G80 was used.
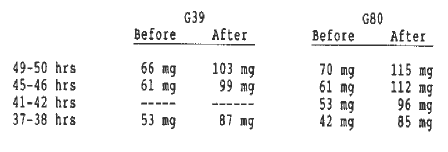
Sample preparation and protein extraction were as described above (Higginbotham and Smith in this Newsletter), but with the following exceptions. Three samples of G39 were prepared with total imbibition times of 37-38, 45-46, and 49-50 hrs. Four samples of G80 were prepared with total imbibition times of 37-38, 41-42, 45-46, and 49-50 hrs. All embryos were labeled during the last 17 hrs of their imbibition period. Each of the seven samples consisted of three embryos. The average mass of the embryos before and after labeling is given in Table 1.
Table 1. Average mass of embryos before and after labeling with 35-S methionine.
The laboratory parameters were as described above (Higginbotham, Smith, and Smith in this Newsletter), but with the following exceptions. A 10.0% SDS-polyacrylamide gel was used in the second dimension. Each of the seven gels that were run (3 G39 and 4 G80) were exposed to film for two periods of time. This generated a light and medium exposure for each gel. The sample of B73 used in the previous study (Higginbotham, Smith, and Smith in this Newsletter) was also run under these conditions and all eight gels were analyzed together.
The data set consisted of 729 spots matched across all eight gels. Of these, 112 spots had zero density values for at least six gels and were deleted from the data set.
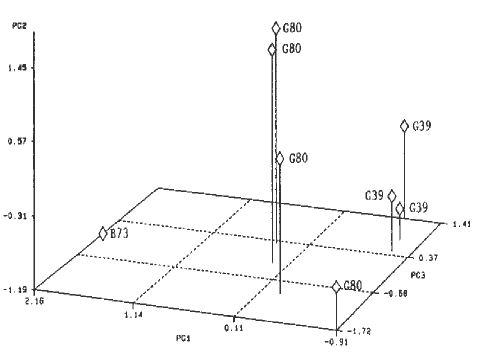
The data were then subjected to principal component analysis with no further spot selection. Figure 1 shows the placement of the eight gels on the first three vectors. The first three vectors encompassed 27%, 25%, and 16% of the variation respectively. Even with slightly varying imbibition periods, principal component analysis succeeded in distinguishing gels of the different inbreds.
Figure 1. Placement of eight gels on the first three vectors of a principal component analysis.
A set of spots was generated which included only those spots where at least 80% of their variation was partitioned between the three inbreds. This set contained 82 spots (13.3%). Another set was generated which included only those spots which had a pattern of variation indicating they may be under regulatory control during this time interval. This set included 40 spots (6.5%). The two subsets shared 10 spots.
The set of 82 spots was subsequently subjected to cluster analysis to ascertain the relative similarity of the inbred protein profiles. G39 and G80 are more similar to each other than either is to B73. The same relative similarity between these three inbreds is obtained with pedigree data.
Among the causes of variability studied in our lab slight differences in the relative development of the seedlings seem to introduce more variation than any other factor save differences in genotype. Moreover some spots which are regulated during this stage of the life cycle may also vary among genotypes. It appears, though, that the set of proteins which are informative of genetic associations will always be large enough to delete those which appear to be developmentally regulated provided those proteins are known.
Finally, it appears that two-dimensional protein profiles will be useful
in determining genetic associations among inbred genotypes. There are compelling
reasons for using this technique in genetic and numerical taxonomic studies.
Given good quality gels and fluorographs these complex molecular profiles
appear extraordinarily stable and reproducible. The effects of different
genotypes on the profile are obvious and measurable. This technique samples
a greater portion of the total genome than any other technique currently
available with the possible exception of the restriction fragment length
polymorphism technique. Not only are polymorphisms at structural loci revealed,
polymorphisms at regulatory loci are also revealed (through quantitative
differences). The data appear amenable to traditional methods of numerical
taxonomic analysis. Additional studies utilizing two-dimensional protein
profiles are in progress in Johnston and Cold Spring Harbor.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}