--Philip S. Stinard and Donald S. Robertson
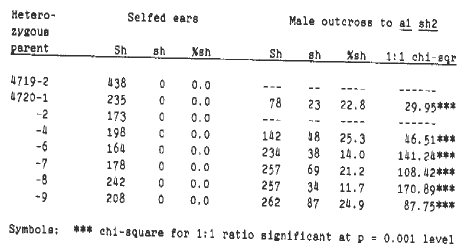
In the 1988 MNL (62:14), we reported a deletion of the A1 and Sh2 loci on the long arm of chromosome 3, a1-sh2-Mu, that arose in our Mutator stocks. In our 1987-88 winter nursery, we planted colorless kernels from the cross a1-sh2-Mu/A1 Sh2 X a1 a1 Sh2 Sh2. We self-pollinated the resulting plants (a1-sh2-Mu/a1 Sh2) in an attempt to obtain a homozygous deletion stock, and outcrossed them to standard a1 sh2 testers to verify the presence of the deletion. Eight selfs were obtained, and of these eight selfed plants, six were successfully outcrossed (Table 1). The reduced frequency of shrunken kernels (less than 50%, p < 0.001) on the outcross ears verified the presence of the deletion in the selfed parents. One of the eight selfed ears segregated for a small, empty pericarp defective kernel mutant, but it is doubtful that this represents the deletion homozygote since seven of the selfed ears had no defective kernels. Had the deletion homozygote been viable, we would have expected a frequency of 0.464 x 0.185 = 0.086 (female transmission x male transmission) shrunken kernels. Since no shrunken kernels were observed, a1-sh2-Mu is most likely homozygous inviable, perhaps even in the zygote.
Table 1. Counts of plump and shrunken kernels on the selfs and outcrosses to a1 sh2 testers of plants heterozygous for the deletion a1-sh2-Mu (a1-sh2-Mu/a1 Sh2).
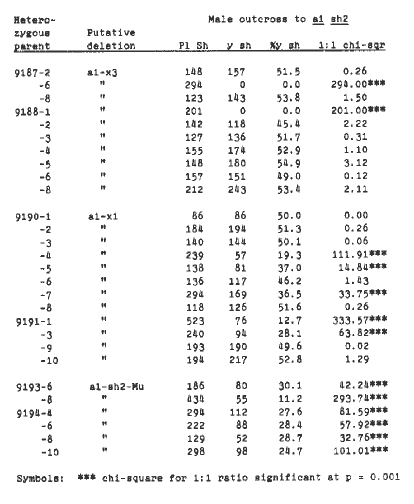
To further characterize this deletion, and to determine the phenotype of hypoploid deletion embryo and endosperm in the presence of hyperploid endosperm and embryo respectively, we crossed deletion heterozygotes (a1-sh2-Mu/A1 Sh2) by TB-3La (a1 3, 3-B, A1 B-3, A1 B-3). Plants heterozygous for the x-ray induced deletions a1-x1 and a1-x3 (L. J. Stadler and H. Roman, Genetics 33:273-303, 1948; both deletions are homozygous lethal) were also crossed by TB-3La for purposes of comparison. All plants were also outcrossed as males to standard a1 sh2 testers in order to determine which plants carried deletions. (Plants grown from the a1-sh2-Mu deletion stock were either a1-sh2-Mu/A1 Sh2 or A1 Sh2/A1 Sh2, and plants grown from the a1-x1 and a1-x3 deletion stocks were either a1-x/A1 Sh2 or a1 sh2/A1 Sh2.) Plants heterozygous for a deletion are expected to have reduced male transmission of the deletion, resulting in fewer than 50% shrunken kernels on the male outcross ear.
The results of the testcrosses to a1 sh2 are presented in Table 2. All outcross ears showing a significant reduction from 50% in the number of yellow shrunken kernels (p < 0.05) were presumed to be outcrosses of deletion heterozygotes. Plants heterozygous for the a1-x3 deletion showed no male transmission of the deletion, which is consistent with the data of Stadler and Roman. The percentage of yellow shrunken kernels on the outcross ears of a1-x1 and a1-sh2-Mu deletion heterozygotes varied from 12.7% to 37.0%, and from 11.2% to 30.1% respectively, which is also consistent with previous data. Kernel counts for the outcross of A1 Sh2/A1 Sh2 sibling plants of the a1-sh2-Mu heterozygotes are omitted from this table, but gave 100% plump purple kernels, as expected.
Table 2. Counts of purple plump and yellow shrunken kernels from the male outcrosses to a1 sh2 testers of plants heterozygous for either the standard a1 sh2 alleles (A1 Sh2/a1 sh2) or various a1 sh2 deletions (A1 Sh2/a1-sh2-del).
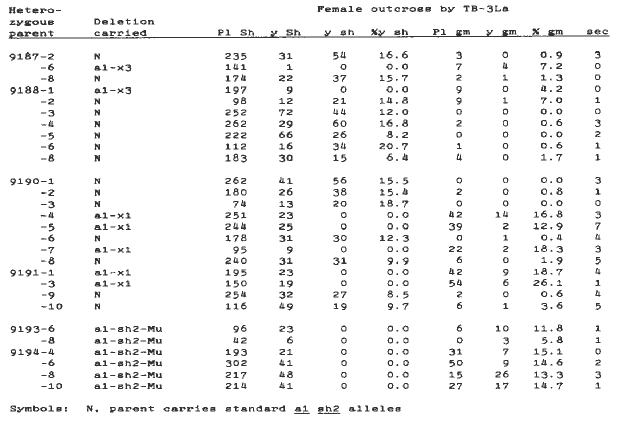
Kernel counts from the crosses by TB-3La of the heterozygous deletion plants (a1-del/A1 Sh2) and their normal siblings (a1 sh2/A1 Sh2) are presented in Table 3. In addition to the expected purple plump kernels, the normal plants segregated for yellow shrunken kernels (a1 a1 sh2 sh2 3-B hypoploid endosperm, with a1 sh2, 3-B, A1 B-3, A1 B-3 hyperploid embryo) and yellow plump kernels (a1 a1 a1 sh2 sh2 Sh2 endosperm from pollination of a1 a1 sh2 sh2 endosperm polar nuclei by the normal a1 3 homologue carried by the TB stock). A small number of germless kernels (under 2% in most instances) were also observed, probably induced by drought and heat stress on developing kernels, since germless kernels have not been reported by other workers making this cross.
Crosses of all three heterozygous deletions by TB-3La did not uncover a yellow shrunken phenotype as would be expected if the deletions were viable in hypoploid endosperm. Instead, the following exceptional phenotypes (and presumed genotypes) were observed: (1) yellow plump (a1-del a1-del a1 Sh2 endosperm from the pollination of a1-del a1-del endosperm polar nuclei by the normal a1 3 homologue carried by the TB stock), (2) purple germless (a1-del a1-del, 3-B, A1 B-3, A1 B-3 hyperploid endosperm, with an inviable a1-del 3-B hypoploid embryo, and (3) yellow germless (a1-del a1-del, 3-B, a1 B-3, a1 B-3 hyperploid endosperm resulting from a crossover between a1 and the B-3 breakpoint in the TB-3La parent, with an inviable a1-del 3-B hypoploid embryo). The frequency of germless kernels was significantly greater in the crosses of the deletion heterozygotes by TB-3La than in the crosses of the normal sibling plants by TB-3La for all three deletions, so we presume that the excess of germless kernels in the crosses involving the deletion heterozygotes represents lethality of these deletions in hypoploid embryos.
Table 3. Counts of purple plump, yellow plump, yellow shrunken, purple germless, yellow germless, and sectored kernels on female outcrosses by TB-3La (a1 3, 3-B, A1 B-3, A1 B-3) of the heterozygous parent plants listed in Table 2.
If these deletions are lethal in the hypoploid condition in both embryo and endosperm, we might expect to find a class of kernels complementary to the plump germless kernels, i.e., kernels with defective hypoploid endosperm but normal appearing hyperploid germs. It seems from the absence of this class that such kernels abort early in development. We did observe defective kernels on ears crossed by TB-3La, but these did not have germs, and appeared on both the deletion heterozygote ears and their normal siblings; these defective kernels may be drought induced, or perhaps the missing class of hypoploid deletion endosperm kernels is of similar phenotype to kernels aborted by drought (mostly empty pericarp). If the latter is true, then the hyperploid embryo does not develop in the presence of hypoploid deletion endosperm.
The endosperm sectored kernels that occur in rare frequency on the ears
crossed by TB-3La (Table 3) provide some insight into the phenotype of
hypoploid deletion endosperm. These kernels appear to be the result of
chromosome breakage or loss during the development of a1-del a1-del
A1 Sh2 endosperms (B chromosome constitution unknown), resulting in
sectorial hypoploidy for the deletion. When such sectoring occurs on the
crosses of heterozygous normal siblings (a1 sh2/A1 Sh2) by TB-3La,
plump kernels with sectors of yellow shrunken tissue result. When the sectoring
occurs on the crosses of deletion heterozygotes by TB-3La, the sectors
fall into two classes: (1) small sectors, generally less than one fourth
of the kernel area, which are composed entirely of yellow shrunken tissue,
and (2) larger sectors, which have a small border of yellow shrunken tissue
at sector's edge, and collapsed defective endosperm toward the interior
of the sector. (These two types of sectors have been observed in crosses
of a1-x1 and a1-sh2-Mu heterozygotes by TB-3La. They have
not been observed in crosses of a1-x3 heterozygotes by TB-3La, but
the population size for these crosses is small.) Kernels have been observed
that are almost entirely devoid of endosperm except for a small sector
of plump tissue bordered by yellow shrunken tissue; these kernels had small,
but normal appearing embryos. By extension, completely hypoploid endosperms
most likely do not develop, and may not support the development of diploid
or hyperploid embryos, which seem to require the presence of at least a
small amount of normal endosperm tissue in order to develop properly. The
presence of a small border of yellow shrunken tissue at the edge of hypoploid
deletion endosperm sectors indicates that normal tissue adjacent to the
sector is able to provide a diffusible substance that allows bordering
hypoploid deletion tissue to produce an a1 sh2 phenotype rather
than a completely defective phenotype. This substance may be an essential
metabolite produced by a biochemical pathway involving a product coded
by a gene (or genes) which is (are) included in the deleted chromosome
segment. If this is the case, then this metabolite is either not diffusible
from hyperploid endosperm to hypoploid deletion germ, or is unable to overcome
the defect which prevents the development of hypoploid deletion germs.
This model predicts that a defective kernel mutant or mutants (or a zygotic
lethal) will be found on the long arm of chromosome 3 that map to the segments
deleted in a1-x1, a1-x3, and a1-sh2-Mu.
Return to the MNL 63 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}