The Spm element undergoes reversible changes in genetic activity. We have been investigating the genetic and molecular basis of such changes, using active and inactive derivatives of the a-m2 allele of the a locus (McClintock, CIWYB 58:390, 1959; 60:421, 1961; and 61:429, 1962). The a-m2-7991A1 allele has a full-length, autonomous Spm element inserted within the promoter region of the a gene. We have isolated derivatives of this allele, designated a-m2-7991A1-i, in which the element has become genetically inactive. The heritability of the inactive phase differs in the main stalk and the tillers. An inactive element remains largely inactive in the main stalk, although reactivation is sometimes observed more in the second ear than in the first ear. Reactivation of the element is frequently observed in tiller ears. An active element undergoes inactivation more frequently when transmitted through the male parent than through the female parent. These observations suggest that developmental signals in the plant affect the reversal of element activity. Moreover, an active Spm element activates an inactive element (McClintock, CIWYB 58:390,1959; Masson et al., Genetics 177:117, 1987).
We have analyzed the patterns of methylation of the Spm element in plants with active and inactive elements. This was done by digesting genomic DNA with methylation-sensitive restriction enzymes having sites within the Spm element and the flanking a locus. Cleavage at these sites was then assessed by Southern blot hybridization analysis using both a-locus and Spm sequences as probes. Fourteen restriction sites within the element and 7 sites within the a locus were examined.
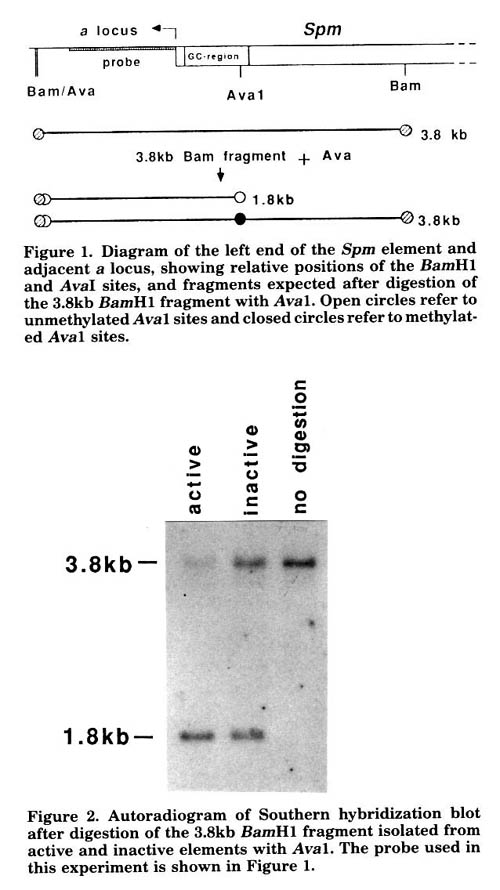
The results of these experiments can be summarized as follows: 1) the a locus sequences flanking the element are always unmethylated regardless of the activity of the Spm element; 2) the Spm element is always methylated regardless of its activity, with the exception of a region of about 600bp at the 5' end of the element, which contains the transcription start site (Pereira et al., EMBO J. 5:835, 1986) and is extremely rich in G and C residues (subsequently referred to as the GC-rich region); 3) methylation of the GC-rich region varies with the element's activity. Within the GC-rich region, an active Spm element is hypomethylated, while an inactive element contains both methylated and unmethylated sites. Furthermore, the ratio of methylated to unmethylated sequences for a given restriction site varies among plants grown from kernels displaying an inactive Spm phenotype. An illustration of the differences in methylation pattern between the active and inactive elements is given here. Genomic DNA from plants heterozygous for the a-m2-7991A1 allele with either an active or inactive Spm and the a allele was first digested with BamH1, an enzyme which is not sensitive to methylation. The 3.8kb BamH1 fragments were then purified from agarose gels following gel electrophoresis. The isolated fragments include the BamH1 fragment containing the element's left end and a locus sequences adjacent to it (Figure 1). The purified fragments were then digested with AvaI, which is sensitive to methylation, fractionated on agarose gels and probed with an a locus fragment (see Figure 1 for location of probe). The 3.8kb BamH1 fragment from the active Spm element is almost completely digested by AvaI, while the fragment from the inactive Spm element is only partially digested by AvaI (Figure 2). Near-complete digestion of the 3.8kb BamH1 fragment by AvaI (and all other methylation-sensitive enzymes with sites within the GC-rich region) is observed in plants with either active elements, spontaneously reactivated elements, or elements reactivated in the presence of other active Spm elements (data not shown).
These genetic and molecular experiments suggest that 1) changes in the activity of the element have no effect on the a locus, which is always unmethylated; 2) methylation of most of the Spm element does not affect its activity; 3) changes in the activity of the element are associated with changes in methylation of a short region at the element's 5' end, which has a very high GC content; 4) an active element must be largely unmethylated within the GC-rich region; and 5) reversal in phase of activity from inactive to active results in demethylation of the GC-rich region.
Figure 1. Diagram of the left end of the Spm element and adjacent a locus, showing relative positions of the BamH1 and AvaI sites, and fragments expected after digestion of the 3.8kb BamH1 fragment with AvaI. Open circles refer to unmethylated AvaI sites and closed circles refer to methylated AvaI sites.
Figure 2. Autoradiogram of Southern hybridization blot after digestion of the 3.8kb BamH1 fragment isolated from active and inactive elements with AvaI. The probe used in this experiment is shown in Figure 1.
J. Banks and N. Fedoroff
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}