In the 1985 and 1987 News Letters (MNL 59:14-15, 1985; 61:10-11, 1987), we presented evidence suggesting that Mutator (Mu) was producing mutations in the male gametophyte. This evidence came from reciprocal crosses of homozygous Y1 Wx Gl1 Mu and y1 y1 wx wx gl1 gl1 non-Mu plants. In such crosses, significantly more white kernels were found when the Mu parent was crossed as a male than when Mu plants were crossed as females. In addition, a significant portion of the kernels from the crosses of Mu plants as males were discordant (i.e., endosperm y1 y1 y1/embryo Y1 y1) whereas when Mu plants were the female parent, few if any discordant kernels were observed. The discordant kernels were suggested to be the result of a mutation in the male gametophyte in which one sperm carried the mutant y1 -Mu allele while the other sperm carried a nonmutated Y1 allele. The discordant seeds resulted when the y1 -Mu sperm fertilized the polar nuclei and the Y1 sperm the egg. Because of the manner in which the female gametophyte develops, this class of discordant seeds is expected rarely if at all as a result of mutations occurring in the gametophyte.
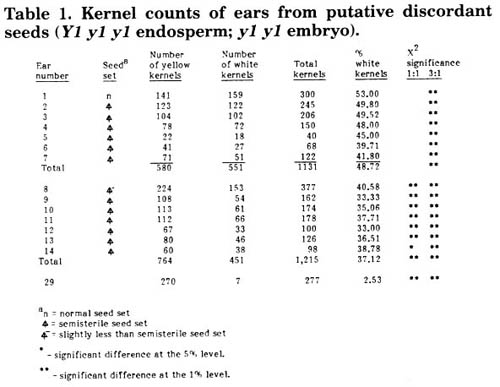
If the discordant seeds are due to mutations in the male gametophyte, there should be a second class of discordant kernels when Mu is used as a male parent (i.e., Y1 y1 y1 endosperm: y1 y1-Mu embryo). Unfortunately this class of discordant kernels, if it occurs, cannot be recognized phenotypically. However, such discordant kernels should give rise to plants which, when self pollinated, will have homozygous white endosperm ears. Thus to determine if such kernels occur, large numbers of yellow kernels from the y1 y1 wx wx gl1 gl1 x Y1 Y1 Wx Wx Gl1 Gl1 Mu crosses must be sown, grown to maturity and the ears scored. Last summer a one acre isolation plot was sown with such kernels and the resulting plants allowed to open-pollinate. In this field half the pollen would carry Y1 and half y1. Thus plants from discordant yellow kernels (i.e., with the y1 y1 -Mu embryos) would have a 1:1 ratio for yellow versus white kernels. Most plants, which are expected not to be discordant, would have a 3:1 ratio of yellow:white kernels. Ears were scored on 37,975 plants and those that differed markedly from the expected 3:1 ratio were harvested and kernel counts made (Table 1). Half of the putative 1:1 ears were in reality not 1:1 but had on average 37% white kernels. These ears had a semisterile seed-set pattern, indicating that perhaps an ovule lethal mutant was induced on chromosome 6 approximately 28 map units removed from the y1 locus. Six ears had 1:1 ratios but had semi-sterile seed set patterns. The plants bearing these ears could have come from discordant kernels which carried some unlinked factor responsible for 50% ovule abortion (e.g., translocation), or they may have had an ovule lethal tightly linked to Y1. Only one ear had the normal seed set and 50% white kernels expected from a yellow discordant kernel. If the plant that bore this latter ear came from a yellow discordant kernel, the frequency of this class of discordant kernels was 2.63 x 10-5. If all of the 7 ears listed in Table 1 were borne on plants from discordant kernels, the frequency of such kernels was 18.43 x 10-5. In last year's News Letter report (MNL 61:10-11), 34.29% of the white endosperm mutant kernels were found to be discordant. If this is the true frequency of this class of discordant kernels among white mutant kernels, 46 of the 135 kernels found in the test summarized in last year's Table 1 would be expected to be discordant. Thus the frequency of white-endosperm discordant seeds in the 275,623 kernel population scored (1987, Table 1) would be 16.69 x 10-5. Because the yellow kernels tested this last summer came from the same population analyzed in the 1987 report (Table 1), it is possible to compare the frequencies of the two classes of the discordant seeds. If only the 1:1 ear with normal seed set is from a discordant kernel, the frequency of the two classes of discordant seeds would be significantly different at the 1% level. However, if all 7 of the 1:1 ears are from discordant kernels there is no significant difference in the frequency of the two kinds of discordant kernels. These results tentatively support the hypothesis of Mu activity in the male gametophyte. However, the 1:1 ears, especially the semisterile ears, could be the result of other phenomena such as a linked ovule lethal. Fortunately, it is possible to test for the basis for the 1:1 ears. As reported last year 71.94% of the Mu-induced y1 mutants have pastel seedlings. If self-pollinated plants from half of the yellow kernels from the 1:1 ears segregate for white kernels, all of which produce pastel seedlings, it would be reasonable to assume that the plants with 1:1 ears came from discordant kernels with embryos of the genotype y1 y1 -Mu. If the 1:1 ratio is due to an ovule abortion gene linked to Y1 most, if not all, of the kernels from these ears should give rise to plants with normal seed set. Since these ears were open-pollinated, a few kernels would be expected to have been produced by pollen from the same plant. Some of this pollen should carry the ovule abortion gene linked to Y1.
It should be noted that Mu-induced mutants in the zygote would give the same results as would mutations in the male gametophyte, and the result considered in this report and the previous ones cannot distinguish between these possibilities.
Table 1. Kernel counts of ears from putative discordant seeds (Y1 y1 y1 endosperm; y1 y1 embryo).
Donald S. Robertson and Philip Stinard
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}