Few, if any, dominant mutations induced by controlling elements have been reported in the literature, and to date none have been characterized molecularly. A dominant virescent mutable mutant has been reported by Peterson, but it apparently had no interaction with the En (Spm) system, in which stocks it arose. In addition, a dominant dappled aleurone mutant arising in a Mutator population has been described by Stinard and Robertson (MNL 61:7), but has not yet been proven to have been induced by the Mutator system. In this paper we describe a dominant amylose-extender mutant which arose in a Mutator population. If this mutant proves to have been induced by a Mu insert, it will provide an excellent opportunity to characterize a dominant mutation at the molecular level.
In our 1984 y1 wx gl8/Y1 Wx Gl8 Mu isolation plot, a single glassy kernel was observed in a population of 388,688 kernels. We planted this kernel in 1985, and selfed and outcrossed the resulting plant to a standard starchy line (Standard Q60). The selfed ear segregated for a sugary kernel type in an approximate 1:2 ratio of starchy to sugary kernels. The outcross ear segregated in an approximate 2:1 ratio of starchy to sugary kernels. Thus, it appeared that our mutant (which we designated Su*-5180) was dominant, but was transmitted in a reduced frequency since the ratios observed differ significantly (p<0.001) from the expected values of 1:3 and 1:1 in the self and outcross respectively (Table 1). Further tests (described below) showed that Su*-5180 is fully female transmissible, but frequently shows reduced transmission through the male.
We tested Su*-5180 for allelism with su1, su2, du, and ae, since these mutants have a sugary or glassy phenotype similar to that of Su*-5180. Selfs of (Su*-5180/su1), (Su*-5180/su2), and (Su*-5180/du) segregated for fully starchy kernels as well as the parental types and their presumed combinations. Thus, we concluded that Su*-5180 was not allelic to su1, su2, or du. The selfs of (Su*-5180/ae), however, produced only sugary or glassy kernels, suggesting that Su*-5180 is a dominant mutant allele of ae. Backcrosses of (Su*-5180/ae) by ae ae stocks also produced ears with only sugary or glassy kernels, confirming the allelism of Su*-5180 with ae. We now designate our dominant amylose-extender mutant Ae-5180.
The expression of Ae-5180 in our standard backgrounds varies from slightly shrunken and tarnished to wrinkled sugary to brittle. This variability occurs whether Ae-5180 is present in the endosperm in 1, 2, or 3 doses. We have made F1 reciprocal crosses of Ae-5180 to several different inbred lines, and observed that the phenotype of the F1 kernels seems to show a maternal effect. When our standard lines carrying Ae-5180 are used as the female parent in the inbred crosses, the F1 kernels exhibit the variability in phenotype noted above. However, when the inbreds are used as females, the expression of Ae-5180 is consistent for a given inbred, but varies depending on which inbred is the female parent. The inbred B73 shows the most extreme expression, the mutant kernels having a glassy, near-brittle appearance. The inbreds Mo17, Oh43, N25, and A636 show good expression, with mutant kernels that are sugary and slightly wrinkled. The inbreds M14, W22, B37, A632, B76, and H99 show fair expression, with smooth, slightly translucent mutant kernels. Mutant kernels can barely be distinguished in crosses to Tama Flint, having a weakly translucent, frosted appearance. We anticipate that after Ae-5180 has been backcrossed into these inbreds for several generations, the above phenotypes will be consistently expressed.
In order to further characterize Ae-5180, and to generate potentially useful stocks, we conducted linkage tests of Ae-5180 with a series of waxy translocations, and the chromosome 5 linkage markers gl8 and a2. The 3 T5-9 waxy translocations that we selected all showed linkage with Ae-5180 (Table 2). The tightest linkage (p = 13.4 cM) was with T5-9c, which has a breakpoint in the short arm of chromosome 5 close to the centromere. This is not surprising, since ae is on the long arm of chromosome 5, about 15 cM from the centromere. We might have expected even closer linkage of Ae-5180 with wx T5-9(4790), which has a breakpoint at 5L.34, especially since ae had been previously shown by J.N. Jenkins (Ph.D. thesis, Purdue, 1960) to be very close to the T4-5c breakpoint at 5L.27. Instead we found linkage of p = 23.1 cM. Such a high percent recombination with waxy could be at least partially explained by the chromosome 9 breakpoint of wx T5-9(4790), 9L.45, being at least 10 cM, and perhaps as many as 26 cM, from the waxy locus on the chromosome 9 linkage map. Crossovers occurring in the region between wx and the chromosome 9 breakpoint would increase the apparent linkage distance between ae and wx in this particular translocation. It is frequently difficult to correlate linkage maps with cytological maps with any degree of precision. The important point to note is that Ae-5180 does indeed show linkage with translocations involving chromosome 5. One other point of interest is that the parental and crossover classes in the wx translocation linkage date are not well-balanced. There seems to be about an equal number of plump (Ae) and sugary (Ae-5180) kernels in each set of translocation data, but for each translocation there seems to be an excess of Wx kernels over wx kernels.
Our linkage data for Ae-5180 and gl8 (p = 12.5 + 0.8 cM, Table 3) are in close agreement with the value of 11 cM reported on the 1987 linkage map. There appears to be an abundance of starchy (Ae) kernels over sugary (Ae-5180) kernels, but this is due to the lower percent germination of the sugary kernels when we were seedling-testing for gl8. The linkage data for a2 and Ae-5180 (Table 4) yield a value (13.1 + 0.7 cM) that is somewhat less than the reported value for a2 to ae of 22 cM, but is in agreement with a 2-point test made by Jenkins, who found a value of 14 cM for a2 to ae in a backcross test.
We have crossed Ae-5180 into homozygous wx stocks in order to observe whether Ae-5180 shows the same phenotypic interaction with waxy as does the standard ae allele. Kernels that are homozygous for the standard ae allele and homozygous for wx are smaller, more translucent, and have finer wrinkling than kernels that are homozygous for ae in a Wx background. The latter kernels are translucent and slightly wrinkled, but not as extremely so as those in a wx background. This effect is very striking, and requires the ae allele to be homozygous (ae ae ae) in the endosperm. If only one or two doses of ae are present, the kernels will be full and plump in either a wx or a Wx background (in certain wx lines, it is possible to distinguish a dosage effect for ae, but we have not observed this in our lines). Ae-5180, on the other hand, can produce the same effect in only one dose. Ae-5180 Ae Ae wx wx wx kernels (where Ae represents the wildtype starchy allele of amylose-extender) are uniformly small, translucent, and finely wrinkled. Ae-5180 Ae Ae Wx Wx Wx kernels, on the other hand, are usually just slightly smaller in size than wildtype starchy kernels, and show the wider phenotypic range from slightly translucent to wrinkled-sugary to brittle. Thus, in both wx and Wx backgrounds, one dose of Ae-5180 in the endosperm appears to have the same phenotypic effect as 3 doses of the standard ae allele.
An analogous effect can be observed when the Ae-5180 and standard ae alleles are crossed into a homozygous su1 background. It has been reported (H. Kramer et al., Agron. J. 48:170) that ae and su1 have an interaction such that kernels homozygous for both mutants are plump, but highly translucent. This unique phenotype is different from that of either homozygous su1 (wrinkled-sugary) or homozygous ae (slightly translucent, slightly wrinkled). We have produced plants that are homozygous su1, but heterozygous for Ae-5180, and outcrossed them to homozygous su1 testers. The resulting ears segregate 1:1 for wrinkled-sugary kernels (Ae Ae Ae su1 su1 su1) and plump, translucent kernels (Ae-5180 Ae Ae su1 su1 su1). Thus, in a homozygous su1 background, one dose of Ae-5180 in the endosperm seems to have the same phenotypic effect as 3 doses of the standard ae allele. It should be noted that the phenotypic interactions described for Ae-5180 with wx and su1 are based on the physical appearance of the kernels, and have not been studied yet biochemically.
In a homozygous wx background, pollen that carries the ae allele can be readily distinguished from pollen that carries the wildtype Ae allele by a differential iodine staining technique (C.W. Moore and R.G. Creech, Genetics 70:611). This technique relies on overstaining the pollen grains with iodine, which is taken up by amylose but not amylopectin, followed by heat destaining. The Ae wx pollen grains, which contain amylopectin but no amylose, stain red. The ae wx pollen grains, which contain starch that has at least some characteristics of amylose, stain dark blue. Using this technique, we found that pollen from Ae-5180 Ae wx wx plants had both dark blue and red staining pollen grains in a 1:1 ratio. Thus it seems that Ae-5180 wx pollen also stains dark blue, opening up opportunities to use differential staining to study reversion rates of Ae-5180 and to do fine-structure mapping of Ae-5180 with respect to other ae alleles.
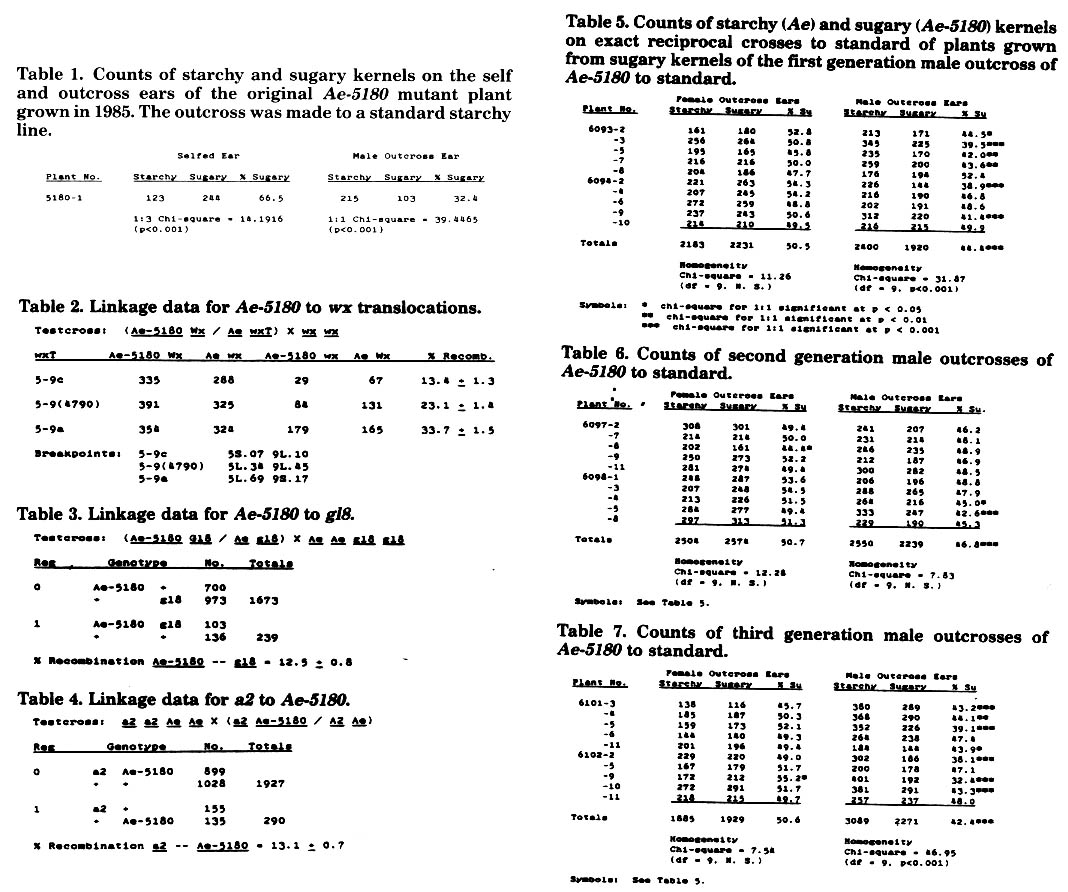
As mentioned previously, Ae-5180 frequently shows a reduced frequency of male transmission in competition with the wildtype Ae allele. In order to study this phenomenon further, we utilized several generations of male outcrosses of heterozygous Ae-5180 Ae plants to standard starchy lines (Ae Ae) in reciprocal crosses with standard lines. All reciprocal crosses were made during the summer of 1987. For the purposes of this discussion, the term "sugary" will be used to describe kernels showing the Ae-5180 phenotype. In all generations, Ae-5180 was fully female transmissible (Tables 5, 6, and 7). However, male transmission varied from generation to generation, and among sibling plants. The data in Table 5 present kernel counts from female and male outcross ears of reciprocally crossed plants grown from sugary kernels of the first generation male outcross ear. None of the male outcross ears showed a transmission of Ae-5180 as low as that of the original outcross ear presented in Table 1 (32.4% mutant kernels), and several ears showed normal transmission.
Table 6 presents kernel counts from second generation male outcross ears. The kernels which were planted to produce family 6097 came from an ear which had 30.8% sugary kernels, and the kernels planted to produce family 6098 came from an ear that had 29.1% sugary kernels. Thus, we expected the plants in families 6097 and 6098 to inherit the system responsible for low male transmission of Ae-5180. As can be seen from the data in Table 6, only 2 of 10 plants showed significantly reduced male transmission of Ae-5180.
Table 7 presents kernel counts from third generation male outcross ears. The kernels planted to produce family 6101 came from an ear with 45.6% sugary kernels. The kernels planted to produce family 6102 came from an ear with 45.8% sugary kernels. Most of the male outcross ears of families 6101 and 6102 showed reduced transmission of Ae-5180, with percentages of sugary kernels comparable to those reported in Table 5. There seems to be no pattern to the reduction in male transmission of Ae-5180 other than that it recurs even after several generations of outcrossing. Some workers (Jenkins, and others) have noted transmission anomalies associated with the standard ae allele. Perhaps there is some inherent biochemical effect of mutant amylose-extender alleles on the ability of mutant pollen to compete with wildtype Ae pollen. The degree to which the competition of mutant pollen is affected might be influenced by genetic background. This would not be without precedent, since wx pollen grains have been observed to germinate more slowly than Wx pollen grains in certain genetic backgrounds, resulting in a lowered frequency of wx transmission. The standard starchy lines that we use in our outcross studies have as their genetic background 4 different inbred lines. Thus, the individual plants in our male transmission studies are fairly heterogeneous. This could account for the wide variation in male transmission that we observe. As mentioned previously, we are in the process of moving Ae-5180 into several different inbred backgrounds in order to control genetic variation in these experiments.
In order to be certain that the reduced male transmission observed in our experiments is a real effect, and not due to poor penetrance of Ae-5180 in some of the heterozygous kernels in the male outcrosses, 100 phenotypically starchy kernels from second-generation male-outcross ears were planted, and the resulting plants selfed. Out of 76 ears obtained, none segregated for sugary kernels. Therefore, the reduced number of sugary kernels in male outcross ears is due to a real reduction in male transmission of Ae-5180, and not due to poor expression of the gene.
In all respects, Ae-5180 behaves as a dominant mutant allele of amylose-extender. This gives rise to a series of questions: (1) How is a dominant mutation responsible for inducing a phenotype previously observed to be induced only by recessive mutant alleles? (2) What is the molecular nature of the wildtype, recessive and dominant amylose-extender alleles, what distinguishes them, and how are they expressed? (3) It has been suggested that Ae is a structural gene that codes for starch branching enzyme IIb (K. Hedman and C.D. Boyer, Biochemical Genetics 20:483). Does the existence of a dominant allele at this locus imply a regulatory role for Ae, or can structural genes be modified to produce a dominant mutant? (4) The Oy locus is another example of a gene that has both dominant and recessive mutant alleles. Will dominant mutant alleles be found for loci previously known to have only recessive mutant alleles (e.g., y1, o2, d1), and conversely, will recessive mutant alleles be found for loci previously known to have only dominant mutant alleles (e.g., Kn, Tu, Pn)? (5) Is Ae-5180 Mutator-induced? We hope that classical and molecular studies of Ae-5180 will provide an opportunity to answer some of these questions. We address the last question in this News Letter.
Table 1. Counts of starchy and sugary kernels on the self and outcross ears of the original Ae-5180 mutant plant grown in 1985. The outcross was made to a standard starchy line.
Table 2. Linkage data for Ae-5180 to wx translocations.
Table 3. Linkage data for Ae-5180 to gl8.
Table 4. Linkage data for a2 to Ae-5180.
Table 5. Counts of starchy (Ae) and sugary (Ae-5180) kernels on exact reciprocal crosses to standard of plants grown from sugary kernels of the first generation male outcross of Ae-5180 to standard.
Table 6. Counts of second generation male outcrosses of Ae-5180 to standard.
Table 7. Counts of third generation male outcrosses of Ae-5180 to standard.
Philip S. Stinard and Donald S. Robertson
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}