In 1983, I reported on the effect of several generations of inbreeding of Mu stocks (Mol. Gen. Genet. 191:86-90, 1983). In this first experiment, different levels of inbreeding were produced by intercrossing between Mu stocks in which the putative level of Mu was doubled each generation for up to 4 generations of inbreeding. In these previous experiments, it was found that Mu activity, as measured by the standard Mu test (i.e., the frequency of seedling mutants transmitted by a Mu plant to its progeny), peaked at the second generation of inbreeding and fell off rapidly thereafter and was lost by the fourth generation of inbreeding. This inbreeding loss phenomenon has been subject to molecular analysis and has been found to be accompanied by modification of Mu1-like transposons.
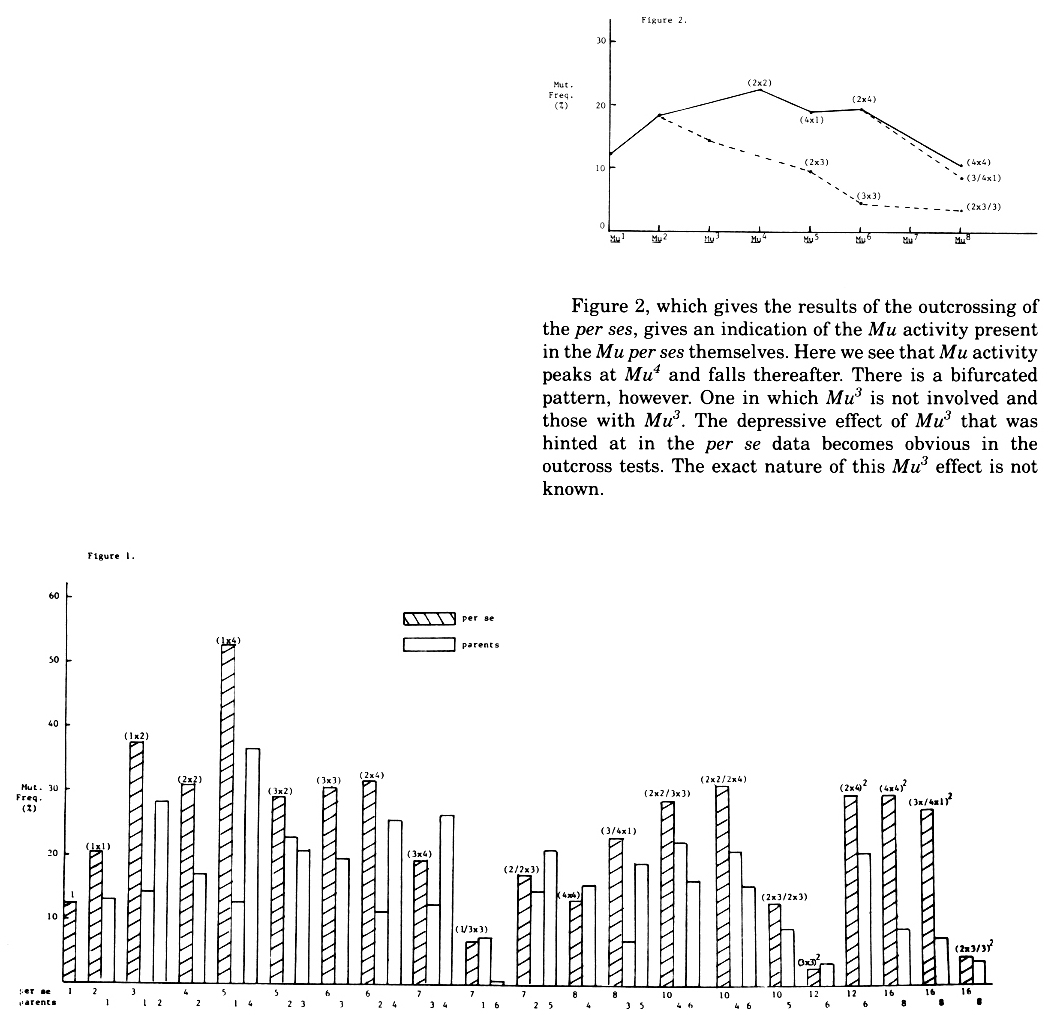
In 1982, we undertook to repeat this inbreeding experiment to determine if the first results were repeatable. We have now obtained the Mu16 per se generation and the Mu8 o.c. (See Table 1 for definitions). In this experiment, we not only made the doubling crosses but also some of the intermediate crosses such as (1 x 3), (1 X 4), (2 x 3), (3 X 4), (3/4 x 1) etc. The results of the per se tests and tests of parents are shown in Figure 1 and the results of the outcross tests of the various per se types are given in Figure 2. We have not yet run a statistical analysis on these results and thus only generalizations can be made at the present time.
Results of the previous experiment, with respect to early generations of inbreeding, are confirmed in that there is an increase in mutants at this level of inbreeding. As inbreeding gets more intense there is a general drop-off of activity exhibited in the per se populations. But there is considerable variation in response depending upon the particular combination of parents. Some more-or-less consistent patterns can be discerned. For example, whenever an Mu3 is involved there tends to be a lower level of activity. The more Mu3 contributes to the per se the greater the reduction in Mu activity; with Mu12 per se (3 x 3)2 showing the least activity. Note also that the straight doubling series (i.e. 1 x 1, 2 x 2, 4 x 4, 8 x 8) behaves as the previous experiment until Mu16 per se. In the previously reported work, there was a sharp drop of activity from Mu8 per se to Mu16 per se. In this present experiment there is instead a sharp increase when going from Mu8 per se to Mu16 per se. We will better be able to determine the significance of this when we can test the Mu activity of Mu16 per se plants by the standard Mu outcross test.
Generally speaking the per ses have more activity than their parents. This is as expected since the per se test is expected to measure the sum of the activity of the parents. However, the per se activity very seldom equals the sum of the individual parent activities or twice the parental activity, as the case may be. The only exceptions are Mu16 per se (4 x 4)2 and Mu16 (3/4 x 1)2. This tendency of the per ses to have fewer mutants than predicted from the parental frequencies suggests either a female effect exhibited by the female Mu parent or a depression of a possible Mu activity that is confined to the zygote or early developmental stages. Such Mu activity has been suggested by previous studies in our laboratory (MNL 60:12-114, 1986).
Figure 2, which gives the results of the outcrossing of the per ses, gives an indication of the Mu activity present in the Mu per ses themselves. Here we see that Mu activity peaks at Mu4 and falls thereafter. There is a bifurcated pattern, however. One in which Mu3 is not involved and those with Mu3. The depressive effect of Mu3 that was hinted at in the per se data becomes obvious in the outcross tests. The exact nature of this Mu3 effect is not known.
The results of this experiment taken together with those from the previous work points up a fact with regard to the Mutator system that cannot be emphasized too strongly. That is, that it is dangerous to generalize from just one experiment or one series of experiments. (For example, one could conclude from the work reported here that Mu3s will be strong inactivators. But we do not know if this response is characteristic of Mu's in general or just the Mu3 in this experiment.) There are probably several reasons for this unpredictability. For example, copy number of Mu elements and state of modification can vary from experiment to experiment and from generation to generation. With just those two variables alone one would not be surprised if two experiments, which superficially appeared to be identical, gave markedly different results. We need to know much more about the Mu system and find some way to standardize our stocks before we can start drawing generalizations from one experiment. This lack of standardization undoubtedly accounts for varying results from different laboratories.
Donald S. Robertson
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}