Taxonomy and phylogeny of the taxa of Zea is still a matter of discussion, and there are completely opposite interpretations despite the large amount of information produced in the last few years. Especially after the last taxonomic treatment of the genus (Doebley and Iltis, 1980) there was produced a wide and diverse body of information that, through morphological, cytological and biochemical studies, has mainly contributed to a correct characterization of the taxa within Zea. Nevertheless, at times, the same available data can lead, depending on the method, to completely different interpretations.
It is because of that, as we have been doing up to now (MNL 59:61, MNL 60:79) and like other authors have also done, we propose an objective handling of the available information through appropriate methodologies, in which each author's preconceived ideas do not have any influence.
Grouping methods by numerical techniques based on a great number of equal importance traits seem to be adequate for this purpose. The objective of this article is to show the results obtained for taxa within Zea, on the basis of 43 traits.
The greatest part of these traits, especially the quantitative ones, were obtained after the evaluation of small populations of the taxa within the genus, and all of them were cultivated under similar environmental conditions. The data obtained from a large number of plants belonging to each of the taxa were used in partial clustering that we have shown before (MNL 59:61; MNL 60:79).
Now, our purpose is to show as a whole those groups carried out on the basis of all our available information. Z. perennis (Zp), Z. diploperennis (Zd), Z. luxurians (Zl), Z. mays ssp. parviglumis var. parviglumis (Zmpp), Z. mays ssp. parviglumis var. huehuetenangensis (Zmph), Z. mays ssp. mexicana (Zmmx) and Z. mays ssp. mays, constitute the 7 operational taxonomic units (OTU's)
Forty-three characters were scored for each OTU as follows: (1) tassel central spike distichous (0), polystichous (1); (2) tassel branch number; (3) tassel branching axis length (cm); (4) tassel central spike length (cm); (5) lateral tassel branch internode length (mm); (6) tassel branching abscission layer absent (0), strongly to weakly developed (1), strongly developed (2); (7) tassel branching rachis flattened (0), rounded (1); (8) distance between the two primary lateral veins of male spikelet outer glume (mm); (9) number of veins between primary lateral veins of male spikelet outer glume; (10) total vein number of male spikelet outer glume; (11) male spikelet outer glume wing absent (0), present (1); (12) male spikelet outer glume primary lateral vein narrow and sparse (0), wide and prominent (1); (13) male spikelet outer glume stiff (1), papery (2); (14) male spikelet outer glume flattened (0), rounded (1); (15) number of fruit cases per spike; (16) female spike distichous (0), polystichous (1); (17) female spikelet arrangement single (1), paired (2); (18) kernel enclosed (1), naked (0); (19) cupule orientation vertical (0), horizontal (1); (20) kernels per cupule one (1), two (2); (21) cupulate fruit case trapezoidal (2), triangular (1), horizontally compressed (0); (22) kernel small (0), intermediate (1), large (2); (23) fruit case weight (mg); (24) fruit case length (mm); (25) fruit case width (mm); (26) fruit case thickness (mm); (27) pericarp thickness (µm); (28) aleurone layer thickness (µm); (29) starch granule size (µm); (30) zein body size of zone 1 (µm); (31) zein body size of zone 2 (µm); (32) zein body size of zone 3 (µm); (33) endosperm protein content (%); (34) Landry-Moureaux saline soluble proteins M; (35) zein; (36) glutelin-1; (37) glutelin-2; (38) glutelin-3; (39) growth habit perennial (1), annual (0); (40) rhizomes present (1), absent (0); (41) chromosome number 2n = 40 (2), 2n = 20 (1); (42) chromosome knob position terminal (1), terminal and internal (2); (43) number of tillers per plant.
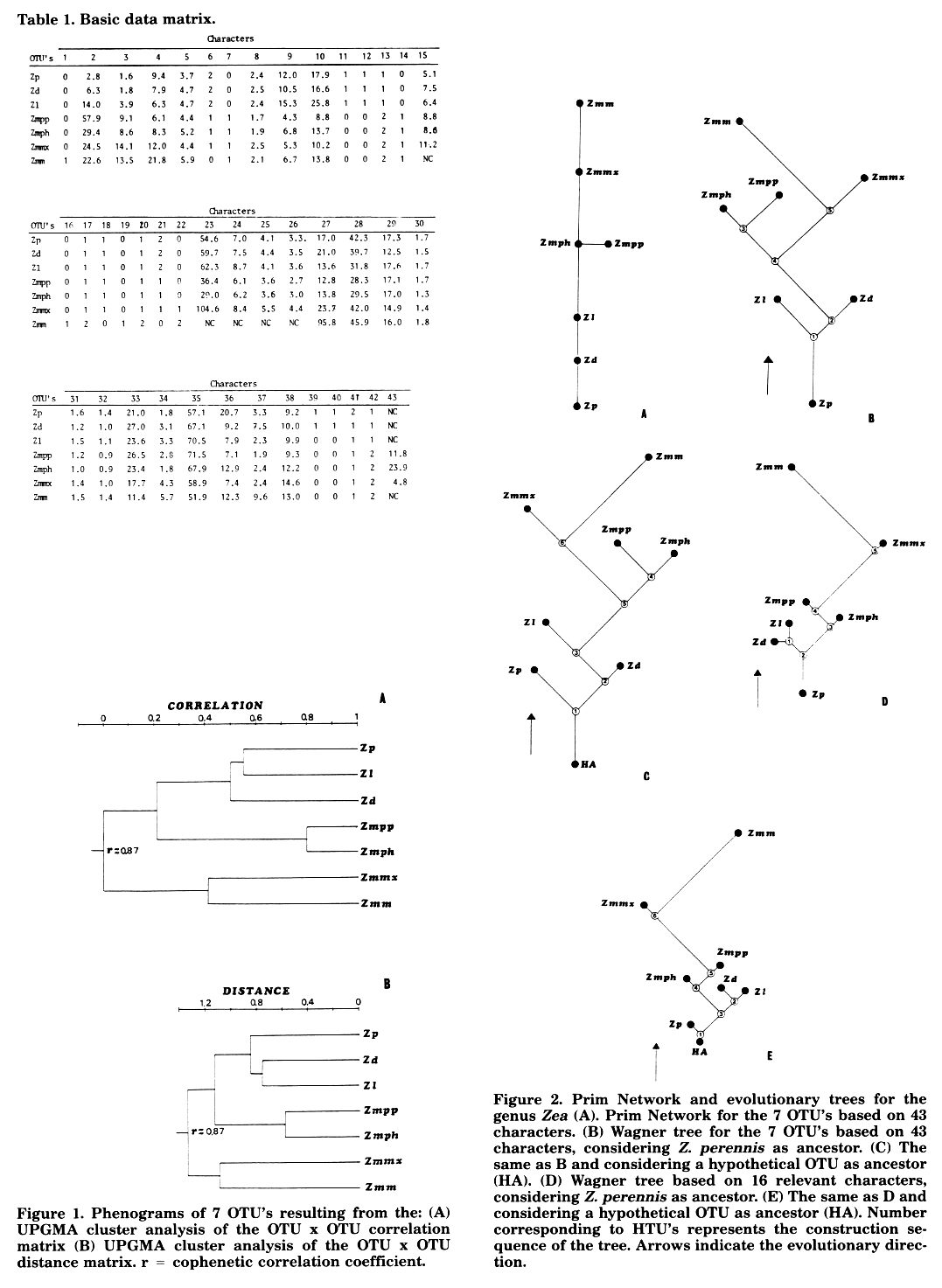
Given a basic data matrix (BDM) (Table 1) of 43 characters by 7 OTU's the data were analyzed by cluster analysis. The BDM was standardized by characters, and phenograms were derived by the Pearson product-moment correlation coefficient applying the unweighted pair group method using arithmetic averages (UPGMA) (Figure 1A) and the "Mean Taxonomic Distance" between pairs of OTU's served as input in the calculation by UPGMA (Figure 113). The "Manhattan Distance" between pairs of OTU's served as input in the calculation of "Prim Network" and "Wagner trees" (Figure 2).
The cluster of taxa, shown in Fig. 1, is partially congruent with those previously carried out (MNL 59:61, MNL 60:79, 60:81) but based on partial information and a smaller amount of data. As can be seen in Fig. 1, as much in one phenogram as in the other a characteristic cluster is repeated, which comes out to be independent of the method employed. Three groups are established, one consisting of Zp-Zd-Zl, which represents the most primitive teosintes of the Luxuriantes section (Doebley and Iltis, 1980). Another group consisting of Zmpp and Zmph is more related to the primitive group of teosintes than to the remaining group, Zmmx-Zmm. It is important to emphasize the fact that different clusters previously carried out (MNL 59:61), and on the basis of different traits, always relate Zp-Zd and Zl in the same group. In contrast, the linkage of the rest of the taxa has been variable, though by now the clusters in Fig. 1 can be considered as much more exact because they are based on a great number of traits.
The taxa of the Zea Section (Doebley and Iltis, 1980) constitute two groups, where Zmpp-Zmph would be much more related to the most primitive teosintes than to the taxa of their own section. This fact states the need of re-examining the taxonomy of section Zea on the basis of appropriate studies. If Zmpp, Zmph and Zmm-Zmmx were variants of the same species they must be linked more closely.
Prim Network and Wagner trees represented in Fig. 2 show a high congruence with the ones previously shown (MNL 59:61; MNL 60:81). The taxa of the genus can be related between two extremes: Zp and Zmm. Just in these two species (perennial teosinte and maize) all the necessary basic information is gathered to produce the other variations shown in the remaining taxa.
Our experience on hybrids between perennial teosinte and maize (see previous MNL issues), with the information that we are considering about the variation in Zea, suggest to us that Zp and Zmm can be considered basic species with which we can experimentally build all (or the greatest part) of the variation in the genus.
Our main idea about the evolution of Zea is like that of other authors, that maize introgression in teosinte (in a primitive teosinte) could have been the main factor in the differentiation of the taxa of this genus. If it were true and could be experimentally demonstrated, the evolutionary trees that we've simulated would be indicating the direction in which introgression increases. The teosintes placed at the bottom of the trees are those which have kept more differences with maize. As we study the top of the trees, the degree of introgression is higher up to a maximum represented by Zmmx, which can be essentially considered a maize which has kept the few necessary genes for wild life.
J.L. Magoja, I.G. Palacios, M.B. Aulicino, L.M. Bertoia and M. Streitenberger
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}