In vitro plant regeneration in maize can be obtained either through organogenesis or by somatic (nonzygotic) embryogenesis. The phenotypic alterations (somaclonal variation) found in regenerated plants have a strong dependence on such morphogenetic events. Quite recently, Rapela (MNL 58:106108, 1984) proposed a model experimental system in maize to obtain organogenesis or somatic embryogenesis working with local materials. The true nature of the morphogenetic events was tested using SEM examination (Rapela and Herkovits, Rev. Fac. Agr. UBA 5:95-105, 1984; MNL 58:108-109, 1984; Rapela, Bol. Asoc. Cien. Nat. Lit. 4:8-13, 1984).
We have obtained several plants regenerated through organogenesis or by somatic embryogenesis. Plants regenerated through organogenesis were aberrant, with several phenotypic alterations, and were very difficult to transfer to soil. Only one of 22 organogenic-regenerated plants reached maturation. On the other hand, plants regenerated through somatic embryogenesis were quite normal and relatively more easy to transfer to soil. The molecular analysis of the association of a particular phenotype with a particular polypeptide configuration was not carried out in organogenic-regenerated plants due to the fact that these plants were always in different development stages. However, among our regenerated plants, 3 floury-a and 5 normal red flint (NRF) plants regenerated through somatic embryogenesis from one single callus each (originated from an immature embryo) and reached maturation almost at the same time (5 days of difference among plants). These plants are adequate for such analysis.
The terminal 10 cm of the upper leaf of such regenerated plants were removed and cut into small pieces. Polypeptides were extracted in two volumes of extraction buffer (200 mM Tris-HCI, pH 8, 5% SDS, 7.5% 2-mercaptoethanol and 20% glycerol), with homogenization in porcelain mortars at 0-5 C. The mixture was allowed to sit on ice for 10 minutes. The homogenization step was repeated and the homogenates were centrifuged at 12,360 g for 10 minutes. The supernatants were transferred to glass tubes, boiled for 5 minutes and stored frozen. Electrophoresis was carried out on 8.3% polyacrylamide gels prepared according to the method of Laemmli (1970). Gels were fixed for 24 hours in 3:1: 1, methanolacetic acid-water, stained in 0.2% Coomassie Brilliant Blue R-250 and destained in 7% acetic acid. Two independent preparations and two gels were run for each plant.
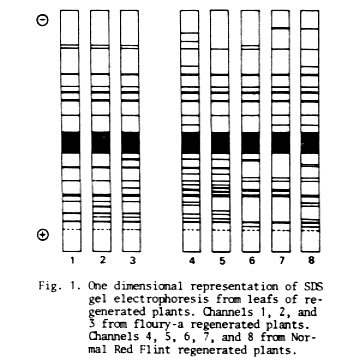
As shown in Figure 1, the SDS-protein pattern of floury-a and NRF maize plants regenerated through somatic embryogenesis from one single-embryo has a particular configuration for each plant. There is a detectable variation between the SDS-polypeptides present in each plant extract and any counterparts.
An analysis based on presence or absence of polypeptides (without quantifying different staining intensities) shows a similarity index (SI = No. of pairs of similar bands / No. of different bands + No. of pairs of similar bands) of 1-2 = 0.88; 1-3 = 0.82; 2-3 = 0.83, for floury-a plants and, 4-5 = 0.59; 4-6 = 0.48; 4-7 = 0.48; 4-8 = 0.52; 5-6 = 0.48; 5-7 = 0.42; 5-8= 0.36; 6-7= 0.75; 6-8= 0.55; 7-8= 0.65, for NRF plants.
Cultures were initiated and the plants were regenerated and transplanted to vermiculite and then to soil at the same time. Therefore, although differences in the SDS-protein pattern due to environmental conditions and/or different developmental stages among plants prior to extraction cannot be ruled out, they are minimized. Eight of the 10 Sl determined for NRF plants were lower than 0.6, indicating a strong qualitative difference between patterns. The percentage of Sl variation due to somaclonal variation (genetic + epigenetic), although unknown under our experimental conditions, is possibly high. Moreover, the SI differences between floury-a (SI average= 0.84) and NRF (SI average= 0.53) plants seem to have a genotypic dependence, and support this last point of view.
Fig. 1. One dimensional representation of SDS gel electrophoresis from leafs of regenerated plants. Channels 1, 2, and 3 from floury-a regenerated plants. Channels 4, 5, 6, 7, and 8 from Normal Red Flint regenerated plants.
Miguel Angel Rapela
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}