Last year we reported on the orientation of the initial divisions of the zygote. Since then we have extended our cell lineage analysis to several later stages of development. Developing embryos of the genotype B/b;Pl/pl;Wd/wd were irradiated (500 R) at various times after pollination. At the time of irradiation samples were taken from a few representative ears and were then examined histologically to determine the developmental stage of the embryo. Mature seeds from irradiated ears were planted and the size and extent of white, green and sun-red sectors were recorded as soon as the sectors became visible. White wd sectors could be scored along the entire height of the plant, although only leaf-sectors could be scored in the case of the lowermost nodes. Sectors of b and pl are not well expressed in either leaves or underground internodes and therefore could only be scored above the 7th or 8th node. The apparent cell number (ACN) of a node was usually determined by dividing the circumference of an internode by the width of the sector within it. The ACN of the nodes below ground was determined from the width of a sector relative to the width of the leaf blade. This approach is not very satisfactory because the leaf blade is not always derived from all the cells in the circumference of the node, and the extent to which a sector expands varies depending upon its position within the leaf. Unfortunately, it is the only practical method available.
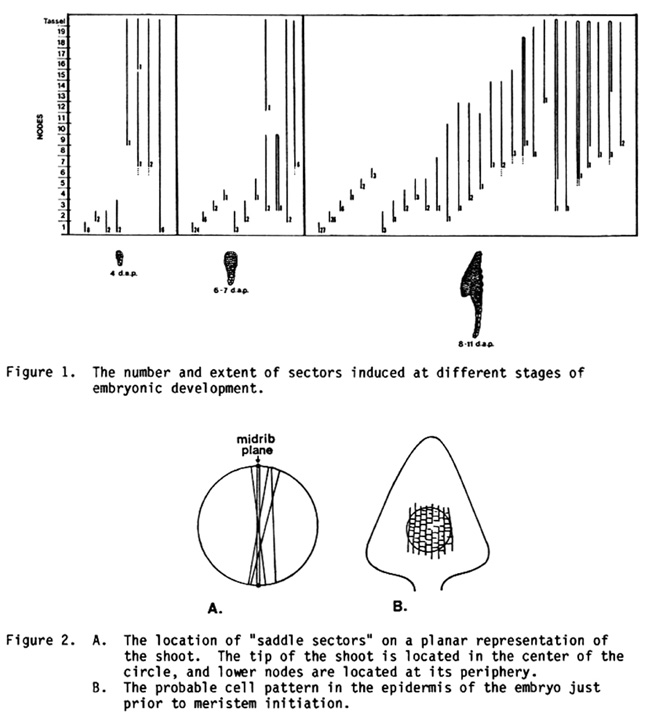
The longitudinal extent of the sectors we observed is illustrated in Figure 1. As we have previously noted, the variability in clonal patterns is quite considerable. Nevertheless, several consistent features emerge. One of the most striking of these is the early separation of the lineages of the first and second nodes from the upper nodes. Four days after pollination many sectors still ran the entire length of the plant; but by the transition stage of development (6-7 DAP), over half of the sectors were restricted to the first node. By the coleoptile stage (8-11 DAP), the second node has also become clonally separated from other nodes. This feature is part of a general process of clonal restriction that proceeds from lower to upper nodes. A similar phenomenon occurs in the shoot apex during germination (Johri and Coe, MNL 54).
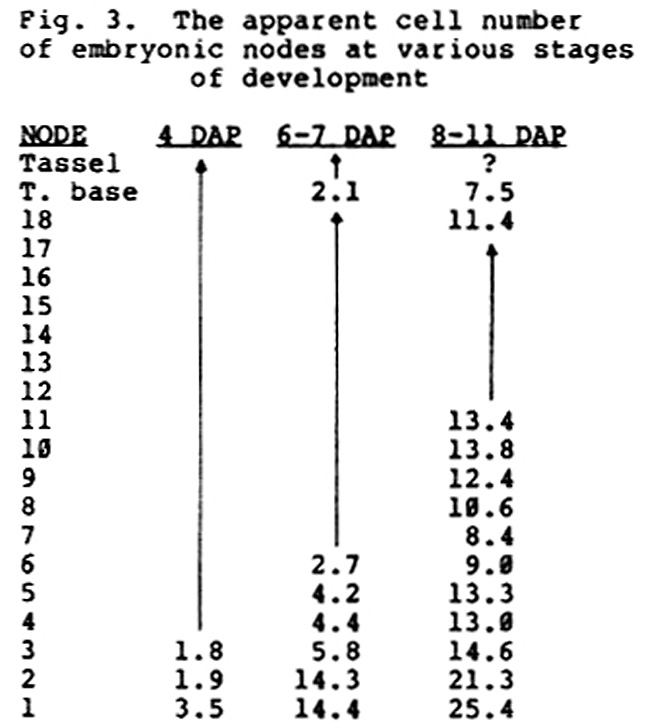
Another interesting feature of these data is the frequent appearance of "saddle sectors," that is, sectors that run from one side of the plant to the other (Figure 2). These sectors are generally restricted to one side of the midrib; on a planar projection of the shoot, they run parallel to the midrib line rather than perpendicular to it. This type of sector indicates that cell division is initially oriented perpendicular to the longitudinal axis of the embryo, so as to produce longitudinal files of cells. If cell division were oriented radially, one would expect saddle sectors to be randomly oriented relative to the longitudinal axis of the embryo (as defined by the plane of the midrib). Thus the apical meristem arises in a region of the embryo that has a highly polarized cell pattern. In this respect, shoot initiation in corn may involve a process similar to that described by Green and Brooks (AJB 65:13-26, 1978) in Graptopetalum.
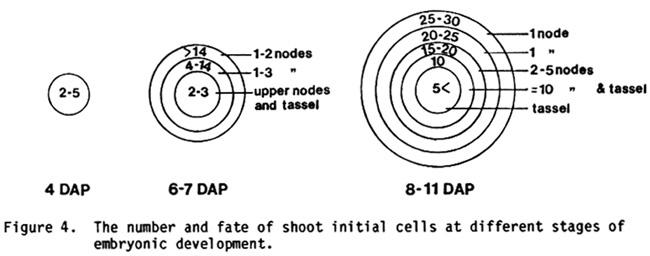
In order to model the shoot apex at various stages in its development it is important to have an estimate of its ACN. Unfortunately, we observed relatively few sectors--particularly at early stages of development--so our estimates are only rough approximations (Figure 3). Nevertheless, they yield a picture that is consistent with the morphology of the embryo. The apical meristem of the shoot arises from a field of cells on the lateral face of the embryo. Since the meristem is circular in cross-section, it is reasonable to model this field as a circle within the paradermal plane of the embryo. It is clear from the size and extent of clones induced prior to the appearance of the apical meristem (i.e., 4 DAP and 6-7 DAP) that this field encompasses a fairly large group of cells, whose members contribute to a limited and more-or-less specific node or set of nodes. On topological grounds, one would expect the lower nodes of the plant to be derived from cells at the outer periphery of the field, and the upper nodes of the plant to be derived from more central cells. If this is true, then the ACN of the lower nodes should be greater than that of the upper nodes, as indeed it is.
Our interpretation of the data presented in Figures 1 and 3 is illustrated in Figure 4. Four days after pollination 2-5 cells in the L1 layer of the embryo are destined to form the shoot apex. Only one or two of these cells contribute to the upper nodes of the plant; the remainder contribute solely to the lowermost one or two nodes. It should be emphasized that cell fates are not fixed at this stage since many sectors extend the entire height of the plant. Whether or not a cell contributes to upper nodes is probably determined by its position within this group and its subsequent pattern of cell division. In a 6- to 7-day-old embryo the primordium of the apex consists of an outer ring of 14 or more cells destined to form the first one or two nodes, another ring of 4-14 cells destined to form the second node or a small group (2-3) of more distal nodes, and 2-4 central cells which form all the upper nodes of the plant. It is worth emphasizing this last point: as late as 6-7 days after pollination all the upper nodes of the plant (3 to tassel) may be represented by only 2 cells in the embryo. By the time the shoot meristem is initiated, the population of initial cells has grown even more, and cell fates have become even more restricted. Between 8 and 11 days after pollination this population consists of around 5 rings of cells containing between 50 and 100 cells. The outer 2 rings contribute to the first and second nodes, the third ring forms nodes 3 to 7, the fourth ring, nodes 7 to the base of the tassel, and the central ring forms the tip of the tassel. There are probably 25-30 cells in the circumference of the outermost ring, 20-25 in the second, 15-20 in the third, around 10 in the fourth, and something under 5 in the central-most ring. This last figure is based on the results of clonal analyses of dry seeds, which show the tassel to be derived from around 4 cells. Histological examinations indicate that the meristem of the shoot has about 10 rings of cells in the L1 layer, and about 6 rings in the L2 layer at the coleoptile stage of development. Since most of the sectors we observed were probably in the L2 layer (see Poethig, MNL 57:35), our results appear to accurately represent the size of the meristem at the time of irradiation.
Figure 1. The number and extent of sectors induced at different stages of embryonic development.
Figure
2. A. The location of "saddle sectors" on a planar representation of
the shoot. The tip of the shoot is located in the center of the circle,
and lower nodes are located at its periphery.
B. The probable cell pattern in the epidermis of the embryo just prior
to meristem initiation.
Fig. 3. The apparent cell number of embryonic nodes at various stages of development
Figure 4. The number and fate of shoot initial cells at different stages of embryonic development.
S. Poethig and E. H. Coe, Jr.
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}