The role of the enzyme uridine diphosphate glucose:flavonol 3-0-glucosyltransferase (UFGT) in flavonoid and anthocyanin synthesis has been extensively documented (Larson and Coe, Biochem. Genet 15:153, 1977; Dooner and Nelson, Biochem. Genet. 15:509, 1977). The enzyme acts on the 3 position of flavonoid compounds and is required for anthocyanin stabilization. The purpose of this report is to describe two additional flavonoid-dependent, glucosyltransferases found in extracts of aleurone and endosperm tissues.
Recently we modified the method for isolation and assay of UFGT (Klein and Nelson, PNAS 80:7591). The principal change in our procedure was to replace Tris buffer with Hepes; this improves the stability and yield of UFGT. In the Tris buffer system, no UFGT activity is detectable in preparations from bz1 endosperms (Dooner and Nelson, ibid.; Klein and Nelson, unpublished data). However, with our new protocol, we could now detect low levels of flavonoid-dependent glucosyltransferase activity in bz1 endosperms.
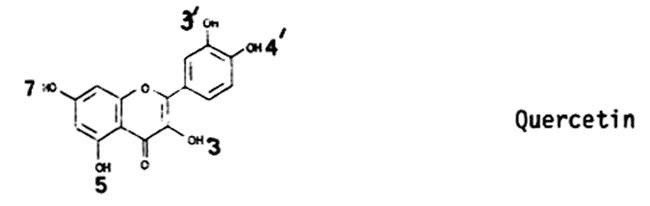
The assay utilizes 14C-labelled UDPglucose and the flavonol quercetin as substrates (Klein and Nelson, ibid.). These minor glucosylated compounds are not resolved from quercetin by paper chromatography in 15% acetic acid; following two dimensional separations two bright yellow, fluorescent spots are visible which are distinct from quercetin, with Rf values as follows:
Even with two dimensional chromatography, resolution of these compounds is poor; this has hampered attempts to isolate enough of each to identify them by spectral properties, etc. It is clear, however, that these represent glucosylation of the flavonol substrate at some position other than the 3 hydroxyl. The latter compound, isoquercitrin, is well separated under these conditions from the unknown compounds.
In order to assess the possible role(s) of the minor flavonoid glucosyltransferases we surveyed three different genotypes for the distribution of these activities. Dr. Coe generously provided the double mutant bz1 bz2.
Both activities (A and B) were found at similar levels in the single bz1 and double mutant bz1 bz2. Thus, neither activity appears to be correlated with the bz2 mutation. Only one radioactive compound, other than isoquercitrin, appeared on chromatograms from assays of Bz1 Bz2 extracts; this had a mobility similar to compound B. The high level of UFGT activity in Bz1 Bz2 extracts may compete with activity A for substrate, reducing the level of product A below the level at which it can be detected. Furthermore it is not clear whether any of these glucosides may be modified at more than one position.
Dooner (Phytochem. 18:749, 1979) described three minor flavonoid-dependent glucosyltransferase activities in extracts prepared from bz1 embryos. One activity appeared to be an isozyme for UFGT; the other two activities produced bright yellow, fluorescent compounds which he identified as the 3'- and 7-glucosides of quercetin. He did not detect these activities in extracts from Bz1 or bz1 endosperms (with aleurone). The two endosperm flavonoid-glucosyltransferase activities we described above produce compounds which have somewhat different mobilities than those described by Dooner. The function of these minor flavonoid-glucosyltransferase activities in vivo has not been established. These enzymatic functions may be involved with the synthesis of six diglucosides of quercetin which Styles and Ceska tentatively identified in Bz pollen (Maydica 26:141, 1981). Some of these compounds were glucosylated at the 3', 4' and/or 7 positions as well as the 3 position of the flavonoid rings.
Anita S. Klein and Oliver E. Nelson, Jr.
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}