There have been only a few studies dealing with different aspects of mitotic analysis of maize, such as banding, somatic association, karyotype vs. translocations, endosperm cytology, etc. The comparatively insignificant work done with the mitotic material is attributable to several technical problems, viz. difficulty of securing uniform one-celled thick squashes in view of the very hard nature of the root tip, and very heavy cytoplasmic content. Filion (MNL 42:175, 1968) further pointed out that maize chromosome arms would not spread very widely, and as such it would be difficult to locate accurately the centromere and to define the limits of arms. In view of the lacunae in our knowledge in understanding the mitotic chromosomes of maize, and also in view of the immense potential of improved methods of chromosome analysis, it was deemed worthwhile to make an attempt to unravel the variation patterns of somatic chromosomes of maize. The present report comprises a detailed investigation of somatic chromosomes of a representative cross-section of the cytological diversity existing in maize (mostly from Indian sources). A cultivar of teosinte and another of Coix lacryma-jobi were included for comparison.
The treatment schedule for mitotic analysis is as follows:
1. Pretreatment: 1:1 mixture of saturated bromonaphthalene and 0.05% colchicine, for 2 hr 45 min at 10-12 C (including three minutes chilling time)
2. Fixation: 1:3 acetic-ethanol overnight
3. Preservation: 70% ethanol
4. Maceration: 5"; aqueous solution of pectinase at 37 C for 2 hr
5. Clearing: 45% acetic acid at room temperature for 10 minutes
6. Staining: 2% aceto-orcein overnight at room temperature
For Feulgen staining, the pretreated, fixed and macerated root tips were hydrolyzed in 1N HCl at 60 C for 15 min, and then stained in Feulgen solution for a period of 2 hr. Alternatively, the maceration might also be carried out after staining, but at room temperature and for a shorter duration.
Based on the nature of primary and secondary constrictions, the chromosomes of the cultivars studied were classified under three types:
A-type: Chromosomes with two constrictions; three subtypes were recognized.
A1-type: Chromosomes with two constrictions, with median to nearly median primary constriction and submedian to nearly submedian secondary constriction.
A2-type: Chromosomes with two constrictions, with median to nearly median primary constriction and subterminal to nearly subterminal secondary constriction.
A3-type: Chromosomes with two constrictions, with submedian to nearly submedian primary constriction and subterminal to nearly subterminal secondary constriction.
B-type: Chromosomes with median to nearly median primary constriction.
C-type: Chromosomes with submedian to nearly submedian primary constriction.
Even though B- and C-type chromosomes would actually form one type, they were classified based on the centromeric index.
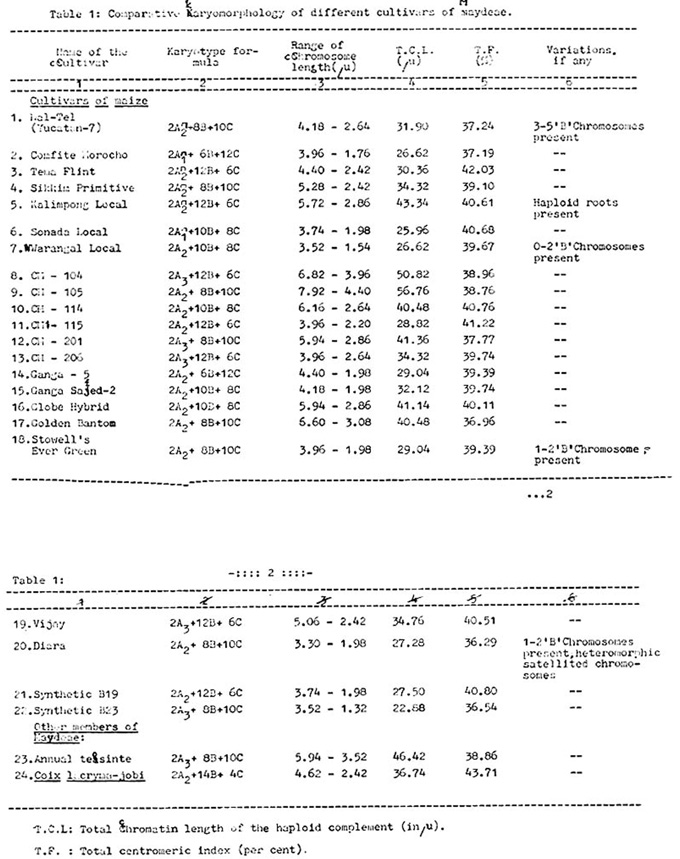
The details of the comparative karyomorphology of the different cultivars studied are presented in Table 1. The karyotype is graded. The qualitative features of the ideograms are relatively uniform. The karyotypes of teosinte and Coix do not differ more than those of cultivars of maize.
The normal somatic chromosome number of all the twenty-four cultivars studied is always twenty, except for occasional increments due to the presence of B-chromosomes in cultivars such as Nal-Tel (Yucatan-7), Warangal Local, Stowell's Evergreen and Diara, which was confirmed by Feulgen staining also. Their number ranges up to a maximum of five, although one or two is the most frequently encountered number. The longest chromosome measures 7.92 u in length, and the shortest chromosome 1.32 u. The total chromatin length of the haploid complements varies from 56.76 u in the case of CM-105, to 22.82 u in the case of Synthetic B23. Maximum TF value is 43.71 in Coix, and the minimum TF value is 36.29 in the cultivar Diara; however, the variation in TF values is relatively less compared to that of the total length of the haploid complements. Chromosomes with secondary constriction are always two in number and A2 is the most frequent type encountered, occurring in sixteen of the twenty-four cultivars studied, followed by A3 and A1 types in six and two cultivars, respectively. More frequently, it is the 6th chromosome in order of length, as is the case within the pachytene complements. The chromosome length within an individual complement ranges gradually from the longest to the shortest chromosome, and no abrupt change in the chromosome length was observed. The features of the ideograms analyzed are more or less uniform. The number of B-type pairs is either 4, 5 or 6, and that of C-type pairs 3, 4, or 5, except in the case of Coix, where there are 7 pairs of B-type and 2 pairs of C-type. Some other variations were also observed, such as the presence of heteromorphic satellited chromosomes in Diara, and the occurrence of haploid roots in Kalimpong Local.
The gross chromosome uniformity of maize is the result of judicious human selection. The presence of gross chromosomal aberrations as the possible cause of the wide range of length variation in the chromosome complements can be ruled out. Addition or deletion of nucleotypic DNA during the formation of the cultivars might possibly have played a role in causing the observed variation. Differential condensation patterns of the chromosomes have also been suggested to result in alterations of chromosome characteristics. In addition, several other factors such as minor aberrations, genetic differences, inherent length variability, nature of the origin of the cultivars and differential response to the treatment, may have acted individually and interacted with each other to produce the wide variation. Thus, mitotic chromosome analysis also brings out significant differences in maize.
Teosinte was regarded as a distinct genus or congeneric or even conspecific with maize. There was also controversy regarding the inclusion of the American and Oriental members in the subtribe Maydeae. The present report does not warrant the segregation of maize and teosinte into distinct genera. The observations further support the inclusion of maize, teosinte and Coix within the Maydeae. (The present investigation was carried out under the supervision of Prof. A. K. Sharma of the Department of Botany, University of Calcutta, Calcutta, India.)
J. S. P. Sarma
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}