The shrunken-1 (Sh1) locus in maize, which codes for the enzyme sucrose synthetase, is of major interest because of its interaction with the Ac Ds controlling element system and its tissue-specific pattern of expression. The determination of the site(s) of integration of Ds in relation to the structural region of Sh1 requires work at the DNA level. In addition, DNA technology will facilitate the study of the tissue-specific pattern of expression of Sh1 since this can be investigated using DNA probes.
A first step towards understanding gene function and expression is the determination of gene structure. Accordingly, we have isolated and characterized Sh1 genomic candidate clones. The candidate Sh1 clones were isolated by screening a Black Mexican corn genomic DNA library contained in the Eco R1 site of the Charon 4A lambdoid bacteriophage vector (Sheldon, 1980, Ph.D. Thesis, University of Wisconsin). To locate candidate Sh1 clones, the DNA library was screened with the insert of the cloned cDNA probe prepared from sucrose synthetase coding mRNA and generously provided to us by Nina Fedoroff. Screening yielded six purified Sh1 candidates from a library of more than 106 clones. Of these six clones, one (17.6) was chosen for further analysis.
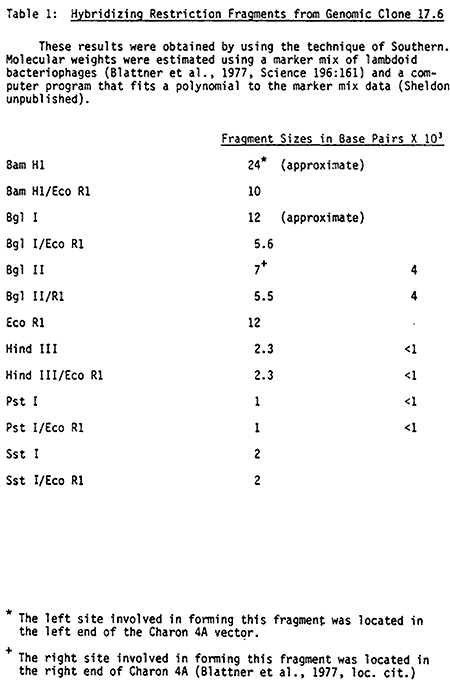
For verification purposes, the candidate Sh1 clone 17.6 was analyzed using the Southern transfer technology. DNA from 17.6 was digested with various restriction enzymes, electrophoresed through agarose, transferred to nitrocellulose, hybridized under our normal conditions at 69 C in 6 X SSC Denhardt's solution to the cDNA probe SS-8 provided by Nina Fedoroff, washed at 69 C in 3 X SSC, and autoradiographed. The results of the Southern analysis are presented in Table 1, which shows the sizes of the fragments from 17.6 which hybridized to SS-8. It is important to note that Bgl II and Pst I each produce two fragments in 17.6 that hybridize to SS-8. Since Bgl II and Pst I each cut the cDNA probe into two fragments (Fedoroff, unpublished and confirmed by us) this result implies that the 17.6 genomic clone and the SS-8 cDNA probe are similar in structure around their Bgl II and Pst I sites. As a further test to verify that 17.6 has regions of good homology with SS-8, hybridization was done under stringent conditions. In this experiment 17.6 DNA was digested with Bgl II, electrophoresed through agarose, transferred to nitrocellulose, hybridized according to our normal procedure, washed in 0.1 X SSC (a stringent condition) and autoradiographed. The same two fragments hybridized to SS-8. This experiment strongly implies that the candidate clone either contains the Sh1 gene or is closely related to it.
We also compared the restriction fragment data from our 17.6 genomic clone to the genomic Southern data generated by Burr and Burr (1981, Genetics, 98:143) and the Starlinger group (Doring et al., Mol. Gen. Genetics, in press). In these comparisons it should be noted that there are at least two important differences. The wild type genotype probed by the above two groups is believed to be the progenitor of some sh1-mutable alleles whereas we used Black Mexican. Secondly, the lengths of the cDNA probes differ. Lengths of 620 and 285 nucleotides were reported by Doring et al. and Burr and Burr, respectively. Fedoroff has estimated the size of the insert in SS-8 to be 700 to 750 nucleotides. From restriction maps of SS-8, using Hind III fragments of pBR 322 as molecular weight markers, we calculate a total length of 700 nucleotides for the SS-8 insert.
In fair accordance with the data of the other groups, we find relatively large Bam H1/Eco R1 and Eco R1 fragments which hybridize to our probe. (The large Bam H1 fragment in Table 1 consists partly of vector.) The smaller size of our Eco R1 fragment may be expected because of the polymorphism in Eco R1 fragments reported by the Burrs. Also in agreement with earlier data, we find a 2 kbp Sst I fragment and a small (less than 1 kbp) Hind III fragment.
Although the Burr and Starlinger groups probed the same genotype, differences in resultant restriction maps were reported. For example, Doring et al. reported an 11.0 kbp Bgl I/Eco R1 fragment but the Burr map predicts a 5.5 kbp Bgl I/Eco R1 fragment. We observed a 5.6 kbp fragment. We do, however, find differences with the Burr map. We place a Bam H1 site within the 5.5 kbp Bgl II/Eco R1 fragment whereas the Burr map shows the Bam H1 site outside of this fragment. This difference in maps could, theoretically, be due to polymorphism.
As mentioned above, analysis of the relatively large Fedoroff cDNA probe reveals internal Pst I and Bgl II sites. As predicted, we observed two bands hybridizing to the probe when the genomic clone was restricted with Pst I or Bgl II (Table 1). Unexpectedly, Hind III digests yield two hybridizing bands. Neither Fedoroff nor we can find an internal Hind III site within the SS-8 insert. It is conceivable that mutational alterations could have created or abolished this internal Hind III site. It is also conceivable that the site of Hind III action is in an intron. Finally, we can not definitely rule out, at this time, the possibility that we have cloned a gene very similar in structure to Sh1.
Edward L. Sheldon and L. Curtis Hannah
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}