Very little evidence is available which allows the identification of gametophyte genetic diversity. Review of the biochemical literature shows that approximately 100 enzymes have been assigned to the tissues of the corn plant but fewer than 40 of these have been identified in mature corn pollen. The techniques of molecular weight polypeptide electrophoresis in concert with computerized digital densitometry (MNL 54, 1980) provide a means by which additional gametophytic genetic diversity can be detected.
Freshly dehiscent pollen was collected from a number of field grown sporophytes and extracted according to the following regime (all operations on ice unless otherwise stated): Buffer added to tissue in the ratio of volume = (tissue fresh weight x 6.89/2 ml). The buffer: 200 mM TRIS-HCl, pH = 7.45; 5% sodium dodecylsulphate; 7.5% 2-mercaptoethanol; and 0.1 M phenylmethylsulphonylfluoride. The pollen/buffer mixture was vortexed and allowed to sit on ice for 20 minutes. The mixture was ground in a motorized tissue homogenizer (15 ml fitted with a teflon pestle), for 1.5 minutes. The homogenate was transferred to a 15 x 100 mm Corex tube and allowed to sit on ice for a further 20 minutes. The homogenate was centrifuged for 20 minutes at 8000 RPM (yields 7.71 Kg with SS-34 rotor on a Sorvall RC-2B centrifuge). The supernatant was decanted and allowed to warm to room temperature following which it was heated for 3 minutes in a boiling water bath, and stored (-20 C) until analyzed. The pellet from the last operation was resuspended in an equal volume of buffer, and stored at -20 C.
Electrophoresis was carried out on homogeneous 8.3% polyacrylamide gels prepared according to the method of Laemmli (1970). Approximately 100 micrograms of polypeptide were applied to each gel well and subjected to an electric field of 80 to 100 milliamp hours. Upon completion of the run, gels were fixed for two hours in 3:1:1, methanol-acetic acid water. Staining was carried out in 2% Coomassie Brilliant Blue R-250. Destaining was accomplished in 7% acetic acid. The stained electropherograms were photographed and subjected to digital densitometry.
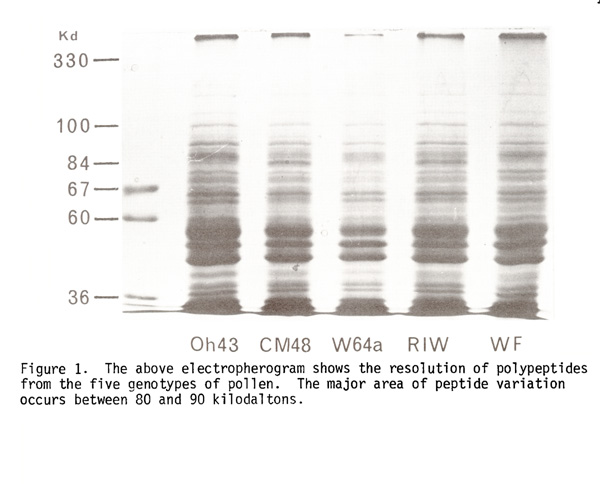
Over one hundred pollen genotypes have been examined employing the above method. Five genotypes were selected as representative of diverse germplasm backgrounds: northern flints (RIW, WF, CM48) and corn belt inbreds (W64A, Oh43) and are described below.
The electropherogram in Figure 1 shows the resolution of polypeptides
from the five genotypes of pollen. The mean number of peptides detected
was 44 per well. In the 80 to 90 kilodalton range of each well, several
peptide variants were detected. Each of the genotypes differ by at least
four peptides within this region. The specific molecular weights (as obtained
by regression and interpolation between standards, mean error 1.6 kilodaltons)
were as follows:
| MW | Oh43 | W64a | CM48 | WF | RIW |
| 89,000 | + | ||||
| 88,742 | + | ||||
| 88,357 | + | ||||
| 87,714 | + | + | |||
| 86,685 | + | ||||
| 86,428 | + | + | |||
| 86,171 | + | ||||
| 85,142 | + | + | |||
| 84,500 | + | + | + | ||
| 84,114 | + | + | |||
| 83,857 | + | ||||
| 82,828 | + | ||||
| 80,642 | + | ||||
| 80,000 | + | + |
The presence of + indicates a particular polypeptide in the corresponding genotype. Note that each genotype differs relative to one another; however, grouping may be assigned according to the derivation of the germ source. The inference of such data is that variant peptides may differ by only a few amino acid constituents. Such variation is regarded as a reflection of genetic diversity in the gametophytic generation.
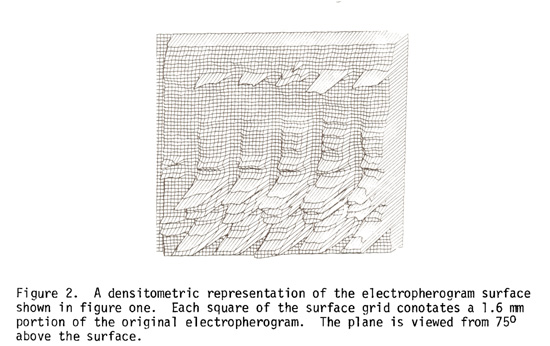
A three-dimensional densitometric "landscape" representation of the electropherogram is presented in Figure 2. The representation was arrived at by averaging 9 x 9 one hundred micron density surface squares to each grid square drawn in the figure. The ratio averaging of density is necessary in order to represent the gel surface in a reasonable size.
Our results from repeated electrophoresis of replicate pollen samples and plantings show very little variation among singular genotypes. This result is not surprising in that many of the stocks employed in the analyses have been subjected to several generations of inbreeding.
Among unrelated lines, electrophoretic analysis has produced consistent band variants (an average of 5-6 bands per genotype). We are now in the process of cataloging such variation (by one- and two-dimensional electrophoresis) in the hope that such data will be of value in characterizing and selecting desirable phenotypic expression. Although our examination of hybrids is only just proceeding, our preliminary results indicate that among intercrosses and F2 generations, segregation of most peptide variants proceeds in a predictable Mendelian fashion. Pollen from a number of plant/plant reciprocal crosses has not yet been processed.
These results demonstrate the feasibility of characterizing polypeptides from pollen. The employment of pollen from aneuploids, aberrations and polyploids will facilitate the opportunity to assign specific peptides to linkage groups. Since these methods can be used to characterize small amounts of tissue, it is now possible to study and compare "landscapes" from germinal and somatic sources by developmental and deterministic methods.
W. G. Hughes and D. B. Walden
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}