Gametophytic expression of many genes has been shown in higher plants (E. Ottaviano and M. Sari Gorla, Israeli-Italian Joint Meeting on Genetics and breeding in Crop Plants, Rome, pp. 89-106, 1979). Direct proof of this phenomenon is obtained when genetical variability is expressed within the pollen population produced by a single heterozygous plant. In maize this has been observed with regard to genes controlling either the reproductive system or sporophytic traits. Some components of the male gametophyte competitive ability are affected by haploid genetic effects. Gametophyte variability resulting from haploid gene expression may play an important role in natural selection, particularly if the phenomenon is related to genes expressed both in sporophytic and gametophytic phase. In maize a positive correlation between pollen competitive ability and values of sporophytic metrical traits has been shown (D. L. Mulcahy, Science 171:1155-1156, 1971 and Nature 249:491-492, 1974). However, in order to evaluate the relative importance of the sporophytic and gametophytic determination of this association, a selection for gametophyte competitive ability experiment was performed, and the correlations between gametophytic and sporophytic traits were evaluated.
The selection for gametophyte competitive ability is based on the different length of the silks from the apex to the ovaries, according to the position of the flower on the ear: the distance decreases from the base flowers upwards. Thus, the probability of fertilization of the most competitive gametophyte, especially if this is due to the pollen tube growth rate, increases from the apex downwards. The Long Ear Synthetic population was used as basic material. From full fertilized ears, two samples of 40 kernels were taken: one from the top and the other from the base. The remaining kernels were discarded. The lines of the first group were reproduced using top kernels, for three selfing generations, while those of the second group were developed using base kernels. The choice of plants for reproduction was random: selection for plant traits was carefully avoided.
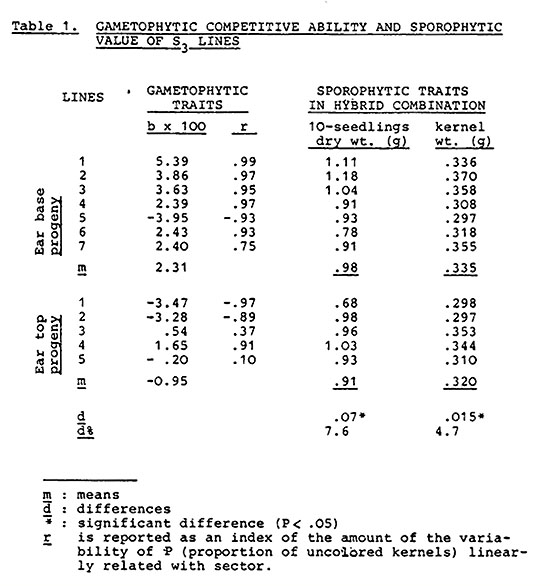
The evaluation of pollen competitive ability was carried out using pollen produced by S3 plants and by means of the pollen mixture technique. Pollen of each line (5 of the first group and 7 of the second) was mixed with an equal volume of pollen of an inbred (W22) with colored aleurone, which was taken as a standard. The mixture was used to pollinate plants of an unrelated F1 (A632 x Mo17) with uncolored aleurone. Ten fully fertilized ears of each line were considered. Each of these was divided into five segments of equal length and scored for aleurone color. The competitive ability, in relation to that of W22, of each selected line was evaluated as variation of the frequency of uncolored kernels from apex to base of the ear. Accordingly, it was estimated as linear regression coefficient of P (proportion of uncolored kernels) on ear segments: a positive value indicates a competitive ability greater than that of the standard. Because the dependent variable is a proportion, referred to binomial data, the method of Armitage was adopted.
Kernel weight and seedling dry weight were considered as sporophytic traits. Kernels from the medium segment of each ear were weighed and sown in the greenhouse for evaluation of seedling weight at two weeks from emergence. For this trial three complete randomized blocks, with plots of ten seedlings, were used.
Table 1 shows the values of the selected S3 lines with regard to gametophyte competitive ability and sporophytic traits in hybrid combination. Positive values of the regression coefficient indicate a gametophyte competitive ability of the selected lines greater than that of the standard (W22). Six of the seven S3 lines obtained from the base of the ear proved to be more competitive than the standard, and one less competitive. A different situation is observed with regard to the lines obtained from apical kernels: competitive ability of S3 lines was less than that of the standard in two cases, in another two about the same, while in the case of only one line was slightly greater. Clearly the choice of kernels from the base or from the apex of the ear led to the formation of two groups of lines differing in male gametophyte competitive value: the long styles of the basal flowers enable the gametophyte with the higher tube growth rate to be selected. The breeding method used operates essentially on within-plant variability, therefore the observed response to selection is a direct proof that haploid expression of genes is an important factor controlling pollen tube growth.
The hybrid progeny of the first S3-line groups (from ear base kernels) proved to have greater kernel and dry seedling mean weights: there was a positive association between differences in gametophyte competitive ability and differences between sporophytic traits. Because gametophyte competitive ability and sporophytic traits were evaluated in separate experiments, the association has to be interpreted as due to genetical causes, arising from linkage and/or pleiotropy. Since the basic material consisted in an open pollinated population, where linkage between sporophytic and gametophytic factors should tend to the equilibrium, it can be concluded that one of the most important factors giving rise to the observed association is the effect of genes expressed both in the haploid and the diploid phase.
E. Ottaviano and M. Sari Gorla
Return to the MNL 54 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}