Allotetraploid maize would be a true-breeding hybrid and would not have the reduction in fertility and vigor caused by aneuploidy in autotetraploid populations.
The allotetraploidization of maize can be achieved by restructuring a maize genome so that its chromosomes will not pair with those of the normal maize genome. This restructuring can be done by concentrating induced and naturally occurring cryptic and visible chromosome aberrations, qualitatively and quantitatively different pairing factors, and genes affecting the expression of differential pairing affinity (like the Ph gene in wheat) into a single line by a recurrent selection type of breeding program.
It has been shown experimentally (Theor. Appl. Genet. 54:103-112, 1979) that differential pairing affinity (DPA) factors are abundant and that they may be readily induced by X-irradiation and chemical mutagens. The assembly of a new genome containing enough DPA factors to produce almost complete autosyndetic pairing presents many problems.
Kind of germ plasm. At one time it was thought desirable to use much exotic germ plasm in the hope of obtaining more naturally occurring DPA factors. This would have the drawback of incorporating much non-adapted material. However, experiments indicate that Corn Belt maize is as variable as that from other parts of the world, and therefore the inclusion of exotic material is not advantageous.
The type of DPA. The DPA could be based on cryptic or visible structural aberrations, or on qualitative or quantitative changes in the pairing code. Inversions are effective in producing DPA, but they pose many problems. They prevent the recombination of genetic material inside the inverted region (except for 4 strand double exchanges) and thus restrict the formation of new combinations of DPA factors. Duplicate-deficient and deficient chromosomes are produced from crossing over in the inverted region. These will persist in tetraploid populations. Reciprocal translocations are not effective in producing DPA according to Sybenga (1973, Genetica 44:270-282). Duplications and deficiencies disrupt the genic balance and are to be avoided.
Thus, it is better to rely on cryptic aberrations; small inversions; and changes in the number of pairing units in the pairing initiation sites if these are repetitive.
Restructuring on the diploid or tetraploid level. It will be done on the tetraploid level for the following reasons. Selection for agronomic traits should be made on the level where the genetic material will be used. The amount of DPA is more easily assessed on the tetraploid level.
Assessment of DPA. Cytological methods of assessing DPA such as determination of quadrivalent frequencies are less efficient than genetic methods. As a plant with the duplex constitution of AAaa approaches allotetraploidy the frequency of aa gametes as determined in a test cross declines from 1/6 (chromosome segregation) to 0. In an allotetraploid only Aa gametes are formed.
A breeding system. The idea expressed by Stebbins and others that an autotetraploid or a segmental allotetraploid can evolve into a true allotetraploid by selection for DPA mutant factors which would produce a more regular meiosis and greater fertility is questionable. Chromosome assortment and crossing over between homoeologues would prevent the fixation of two different pairs of genomes. The basic requirement for speciation is isolation. Consequently, any allotetraploidizing breeding program must keep the population being restructured reproductively isolated from normal maize populations.
This isolation poses a problem. We can grow a population of material which is subjected to X-irradiation and chemical mutagens and take samples from it and cross them with genetic marker stocks. Plants from those crosses which show lower frequencies of recessive segregants (from a higher level of DPA) cannot be used to form an elite population because the restructured material has been mixed with normal material. The use of seed from siblings requires high correlation between it and the material that went into the test cross. Ideally, it should be an inbred line. However, the progress toward homozygosity is very slow in autotetraploid populations (and even slower in segmental allotetraploids)--after 10 generations of self fertilization, the heterozygosity is about 19%. Thus genetic testing would appear not to be feasible.
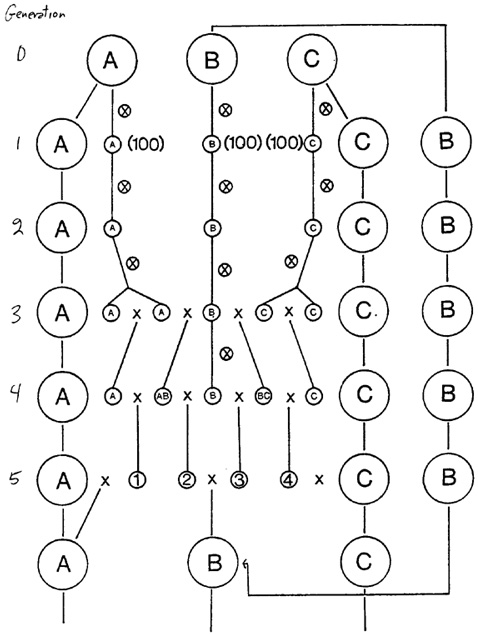
Nevertheless, there is a solution to this problem. It is simply to restructure the genetic markers as well. The breeding system is shown in the diagram.
Three synthetics (A, B, and C) are maintained by random crossing within themselves and are subjected to X-irradiation (10,000 r to the dry kernels) every generation. Synthetic B was derived from 20 diploid maize lines which were subjected to X-irradiation (5,000 r) for 10 generations, 8 exotic lines, various open pollinated varieties which have been incorporated in the tetraploid level via the el gene. This synthetic has a large proportion of Argentine Flint, and Alexander's Synthetic B (a very good source material which was derived from over 30 inbred lines) which were also subjected to recurrent X-irradiation. Synthetic B has all dominant genes.
The other two lines (A and C) are homozygous for a different group of 5 recessive markers: bz2, lg y v16 wx and a su pr gl g. There is one marker for each of the 10 chromosomes.
About 300 plants in each synthetic are grown each year. A random sample of seed from 100 of the best ears is used to produce the next generation. After an initial mixing period (which has been completed) a sample of 100 kernels will be taken to produce test lines. After a few generations of self fertilization, these lines are crossed with each other in the manner shown in the diagram. At generation 4 the test cross is made and the genetic ratios for all ten genes will be determined. A value called the allosyndetic index is computed which is simply the sum of the percentages of recessive gametes. Progenies whose allosyndetic indices are in the 10% lowest group will be selected and backcrossed to the synthetics and added to them. Progenies giving higher allosyndetic indices are discarded.
The ratio of old material to new material in the new synthetic will probably be 1:1. This method carries dominant genes into the A and C synthetics. These can be eliminated easily in future generations. The introduction of recessive genes into the B population may require the use of test crosses in future generations to get rid of them.
Eventually, enough DPA factor will accumulate in the B synthetic, so that samples of it can be taken to be crossed with normal 4n maize lines. It is hoped that one of these hybrids will have 20 pairs of bivalents and be an allotetraploid.
G. G. Doyle
Return to the MNL 54 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}