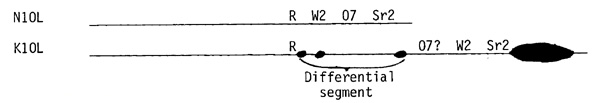
Abnormal chromosome 10 (K10), first described by Longley, differs morphologically from normal 10 in two respects. It is longer by virtue of possessing an extra segment of chromatin at the end of the long arm, whose length at pachynema is approximately equal to that of 10S. This segment contains a proximal euchromatic region, a conspicuous heterochromatic knob, and a short euchromatic tip. Normal 10 not only lacks the extra piece, but differs from K10 in the chromeric structure of the distal one-sixth of its long arm. The corresponding segment in K10 (hereafter referred to as the differential segment) has three small knobs which are not present in N10. The R locus in K10 is situated to the left of the most proximal of the three small knobs. According to Coe's summary, the linear order and map positions of loci in distal 10L are: centromere - R(57) - W2(73) - O7(80) - Sr2(92). These four loci are also carried in the long arm of K10 but aside from the location of R, which is similarly placed in K10 and N10, nothing is known about their order. That the distal one-sixth of N10 differs structurally from K10 was strongly indicated by Kikudome's finding that the 35% recombination between R and Sr2 in N10/N10 homozygotes was reduced to 1% in K10/N10 heterozygotes and that all crossovers between R and Sr2 in K10/N10 plants occurred between R and the leftmost of the three small knobs in the differential segment. It could be argued (1) that the differential segment of K10 is foreign chromatin of unknown origin and that the W2, O7, and Sr2 loci are situated in the portion of K10 extending beyond the tip of N10 or (2) that the differential segment contains the three loci but a rearrangement within the segment reduced crossing over in K10/N10 heterozygotes. On either alternative, the low percentage of R - Sr2 recombination is dependent on a lack of homology between the distal one-sixth of N10 and the corresponding segment of K10. This dissimilarity should be revealed by the pattern of pairing at pachynema in K10/N10 heterozygotes. Somewhat surprising was the observation that pairing of the long arms of the heteromorphic pair was usually intimate, although occasionally asynapsis of the tip of 10L was observed. The data reported in this note demonstrate the correctness of alternative (1) above.
Inasmuch as the K10 chromosome is responsible for the preferential recovery in megasporogenesis of the knobbed chromosome in knobbed/knobless heterozygotes, for neocentromere formation by knobs, and for an enhancement of crossing over in structural heterozygotes and in centric heterochromatin, it is of uncommon cytogenetic interest. We decided to probe the structure of K10 by studying deficient K10 chromosomes resulting from the breakage of dicentric bridges. As we have previously shown in our studies of the high-loss phenomenon, dicentric bridges are produced at the second microspore division because of a delayed replication of the heterochromatic knobs. The K10 knob is similar to knobs on other chromosomes in that it too undergoes delayed replication and chromatin elimination by the above mentioned mechanism. The K10 knob is not completely homologous to other knobs but they all share in common a delayed replication, induced by B chromosomes, at the second microspore mitosis.
When pollen from K10 G R Sr2/K10 G R Sr2 plants with several B chromosomes was used on g r sr2 testers, approximately 80% of the kernels had the anticipated colored aleurone since they received a K10 chromosome carrying the R allele. Sixteen percent of the kernels were colorless having lost the R allele as a consequence of a break in 10L proximal to R and 4% were mosaic for colored and colorless aleurone due to the bridge-breakage-fusion cycle undergone by a chromosome 10 arising from a break distal to R. All of the seedlings from colorless and mosaic kernels were green, indicating the presence of the dominant G and Sr2 alleles in the embryos. However, among the seedlings arising from the colored (R) kernels, some were hemizygous for sr2 or for g sr2 and were phenotypically striate. The g sr2 hemizygotes are grossly deficient for most of 10L and the modified chromosomes were not transmissible through the gametophyte. The hemizygous sr2 plants were tested for the presence of the R allele and for male and female transmissibility of the deficient chromosome. Eleven Df K10 chromosomes have been isolated and are currently being investigated. Our studies are incomplete at this time, but sufficient information has been gathered to permit certain conclusions regarding the structure of K10. The accompanying table summarizes our results to date.
All of the eleven Df K10 chromosomes listed were tested for the extent
of the deficient segment by combining each with strains carrying the recessive
alleles of w2, o7, and sr2. If the F1 aleurone or seedlings expressed the
recessive phenotype, the deficiency was considered to include that locus.
As Brink and McWhirter have shown the recessive opaque-7 trait is subject
to modifying genes and consequently has a low penetrance. It is difficult
at times to know if the apparent normal phenotype is due to presence of
the dominant O7 allele or if O7 has been lost and the pseudo-normal phenotype
results from modifiers. It is, in short, a mutant which should be avoided
if unambiguous results are to be easily obtained (except when the proper
modifiers are present). Since the status of the O7 locus has not been determined
for most of the deficient K10 chromosomes, we have not included it in the
following tabulation.
| Deficient Chromosome |
|
Transmissibility | Cytology | |
| Df K10 (A) | W2 | Sr2 | Female only | Half T* |
| Df K10 (B) | W2 | Sr2 | Female only | Half T |
| Df K10 (C) | W2 | Sr2 | Female only | Unknown |
| Df K10 (D) | Sr2 | Female only | Simple Df? | |
| Df K10 (E) | W2 | Sr2 | Female only | Half T |
| Df K10 (F) | W2 | Sr2 | Female and male | Simple Df |
| Df K10 (G) | W2 | Sr2 | Female only | Half T |
| Df K10 (H) | Sr2 | Female and male | Unknown | |
| Df K10 (I) | W2 | Sr2 | Female only | Unknown |
| Df K10 (J) | W2 | Sr2 | Female, low in male | Unknown |
| Df K10 (K) | Sr2 | Female and male | Unknown | |
*A translocated chromosome involving K10. The complementary translocated chromosome is not present since reciprocal translocations do not occur (Saraiva, 1979).
The Df K10 (F) chromosome has been most extensively studied to date. Df K10 (F) pollen grains achieve considerable success in functioning in competition with N10 pollen with the percentage of functioning Df pollen in different crosses ranging from 18 to 40. The progeny from selfing Df K10 (F) R df/N10 r sr2 plants had, in addition to the expected plump-colored and plump-colorless kernels, an unexpected class with numerous spots of anthocyanin on a colorless background. The spotted phenotype simulated that expected from an unstable r allele mutating to dominant R during endosperm development. Many of the spotted kernels were also mosaics of plump and defective endosperm sectors. The cells with colorless aleurone appeared to coincide with areas having defective endosperm while cells with colored aleurone were over regions with normal endosperm development. The colorless sectors possessed an aleurone layer so the failure of anthocyanin formation cannot be ascribed to absence of the aleurone. The spotted aleurone trait could be accurately classified on most of the ears but considerable variation was found in the degree and extent of defective endosperm development. Some spotted kernels had nearly normal endosperms but the majority were clearly mosaic for normal and abnormal endosperm sectors.
Most of the spotted kernels (R R R) had viable embryos which upon germination gave rise to pure albino seedlings having no trace of green tissue. No albinos came from white (r r r), from mottled (R r r), or from self colored (R R r) kernels, all of which had normal endosperms. In short, the albino seedlings, the failure to synthesize aleurone color, and the abnormal endosperm development were found only when the Df K10 (F) chromosome was homozygous. It may be surmised that the Df K10 (F) chromosome is deficient for a locus involved in regulation of anthocyanin synthesis, for a locus concerned with endosperm development, and for a gene affecting chlorophyll formation, as well as for the Sr2 locus. Presumably separate loci are involved.
Lindstrom (Genetics 1925) stated that the w2 allele, which lies ca. 19 recombination units to the left of Sr2, frequently gave a defective endosperm when homozygous, but this character was sensitive to modifiers since there was great variation in the degree of defectiveness. The twofold effect of the w2 mutation was ascribed by Lindstrom to one pleiotropic or to two closely linked genes. Since the Df K10 (F) chromosome likewise affected both endosperm and chlorophyll, crosses were made of Df K10 (F) R df/N10 r W2 plants with pollen from r w2/r W2 individuals. Self-colored, R-r spotted, and colorless kernels occurred in a ratio of 1:1:2. The plump, self-colored kernels were R df/R df/r W2, the spotted kernels (which were also mosaic for plump and defective endosperm sectors) were R df/R df/ r w2, and the plump colorless kernels were either r W2/r W2/r W2 or r W2/r W2/r w2 in constitution. The self-colored kernels yielded green seedlings, the R - r spotted kernels only white seedlings and the colorless kernels only green seedlings, indicating no or very low crossing over between R and the Df. The Df K10 (F) chromosome was deficient for the W2 locus. Apparently, the w2 chromosome isolated by Lindstrom over 55 years ago has a mutation (or deficiency) for loci controlling aleurone color synthesis as well as endosperm development and chlorophyll synthesis, none of which are complemented by the Df K10 (F) chromosome. The spotted R df/R df/R df kernels from self pollination and the R df/R df/r w2 kernels have identical phenotypes. No homozygous R w2 kernels have as yet been produced in our studies. Our prediction is that they will be phenotypically similar to the above two classes of kernels.
Some of the Df K10 chromosomes do not have a deficiency for the W2 locus. In these cases neither spotted kernels nor white seedlings are produced in F2 populations or in crosses with r W2/r w2 testers.
Although the cytological studies have not been completed, we have found two Df K10 chromosomes lacking the Sr2 and W2 loci which are longer than the normal 10 in Df K10/N10 microsporocytes. Although the Df K10 chromosomes lack the conspicuous K10 knob, they can be distinguished from N10 because they possess the three small knobs in the differential segment. When there is asynapsis of the ends, it is clear that the longer chromosome of the pair is the Df K10. Therefore, the W2 and Sr2 loci must be in the chromatin segment of K10 beyond the end of N10. It should be emphasized that the K10 chromosomes which lack W2 and Sr2 but exceed N10 in length are simple terminal deficiencies. They had little opportunity to accumulate duplicated chromatin by undergoing a series of bridge-breakage-fusion cycles since they arose by breakage of a dicentric bridge at anaphase of the second microspore mitosis. The telophasic products give rise to the two sperm cells. When the sperm nucleus with a freshly broken end unites with the egg, healing of the broken end takes place in the zygotic nucleus.
W2 must lie proximal to the knob since deficient K10 chromosomes have been isolated which are knobless and deficient for Sr2 but still possess W2. Inasmuch as Df K10 chromosomes lacking Sr2 arose from breaks in the proximal euchromatic segment of the extra piece, Sr2 could be in the distal euchromatic tip, in the knob, or in the euchromatin to the right of the breakpoint. A more precise assignment of the cytological location of Sr2 comes from studies by Judith Miles (Indiana University Ph.D. thesis, 1970). She obtained a modified K10 chromosome, here designated Ko, which has the three small knobs of the differential segment and the proximal euchromatin of the extra piece. It was, however, deficient for the knob and the euchromatic tip of K10. Her Ko chromosome had the Sr2 allele. Therefore, the physical location of Sr2 cannot be in the knob or in the distal tip of K10. Since we have chromosomes deficient for Sr2 and doubly deficient for W2 and Sr2 but not for W2 alone, we conclude that the linear order in K10 is centromere - differential segment - W2 - Sr2 - Knob. Although displaced from its normal location, the segment containing W2 and Sr2 has the same orientation relative to the centromere in K10 and N10.
Df K10 (F) may possess the dominant O7 allele, although as stated earlier this trait has a poor penetrance. If this locus has been retained, we have isolated a chromosome deficient for W2 and Sr2 but carrying O7, which lies between these loci. Irrespective of the status of O7, the cytogenetic evidence places the W2 and Sr2 loci in the extra chromatin segment extending beyond the tip of the long arm of N10. If O7 proves to be in the Df K10 (F) chromosome, the linear order in the K10 chromosome has been structurally altered since it would be O7 - W2 - Sr2 - Knob. The origin and constitution of the differential segment has not been established but no genes have been assigned to this segment and our studies indicate that it differs in gene content from the corresponding segment of N10. The differences between N10 and K10 are diagrammatically shown below:
The great reduction in recombination between R and Sr2 in K10/N10 heterozygotes is intelligible by virtue of the transposition of the terminal portion of 10L reported in this note.
M. M. Rhoades and Ellen Dempsey
Return to the MNL 54 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}