Isolation of chemical mutagen-induced oil content mutants of maize
Oil content in maize kernels is almost entirely under genetic control. Evidence for such control is the response of oil content to various selection methods (Sprague and Brimhall, Agron. J. 41:30-33, 1949; Sprague et al., Agron. J. 44:329-331, 1952; Jellum and Marion, Crop Sci. 6:41-42, 1966) and the inheritance of oil content as an independent characteristic (Curtis et al., Agron. J. 48:551-555, 1956; Alexander and Lambert, Crop Sci. 8:272-274, 1968). It appears that from 20 to 40 genes are involved in the determination of oil content in the kernel and these genes exhibit incomplete dominance (Sprague and Brimhall, 1949; Miller, Agron. J. 43:229-234, 1951; Plewa and Weber, Can. J. Genet. Cytol. 15:313-320, 1973). Although many investigators have studied the genetics of oil production in corn, no point mutations that substantially influence oil content have been isolated. I proposed to induce oil content mutations with chemical mutagens. Mutant kernels would be detected and isolated by analysis of oil content with wide-line nuclear magnetic resonance spectroscopy (NMR) (Alexander et al., J. Am. Oil Chem. Soc. 44:555-558, 1967).
The maize lines used in this study were: R805, I-274, R84, IHO, ILO, R804, Alexho 197, Alexho 228, Alexho 724 and Alexho 274. The plants were exposed to the chemical mutagens ethylmethanesulfonate (EMS) and b-propriolactone (BPL). Each maize line was planted at the South Farms at the University of Illinois.
Within each line, pollen from each plant was pooled in a large test tube. The chemical mutagen was prepared in paraffin oil and applied to the pollen grains. The concentration of each mutagen in paraffin oil was 0.13% for EMS and 1% for BPL. The pollen grains were suspended in the mutagen for 20 min. The mutagen-pollen slurry was painted on the corn silks and sibling crosses were made within each line. Each M1 plant was self-crossed and 40 kernels from each M2 cob were individually analyzed by NMR. A representative sample of 10,564 kernels from 265 M2 families were tested for segregation with a Varian PAT-20 NMR.
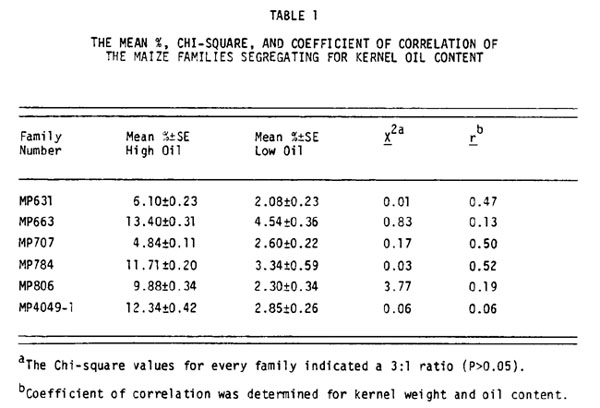
Of the 265 M2 families analyzed for kernel oil content, five families exhibited an increase in variance. These exceptional families were MP631, MP663, MP707, MP784, and MP806. Families MP663, MP784 and MP806 were derived from IHO and families MP631 and MP707 were derived from R84. All exceptional families were from lines treated with EMS. If a recessive allele that affects the oil content of a kernel was induced by a mutagen, a 3:1 ratio of normal to mutant phenotype would appear in the samples from the M2 cobs. The data indicate a bimodal distribution and a 3:1 ratio was indicated for each family after analysis by the Chi-square test (Table 1).
The lower kernel oil content in the presumed mutants could be due to a smaller germ, decreased concentration of oil in the germ, or both. If a mutation that lowered the oil content of a kernel did so by reducing the size of the germ, and if endosperm weight was unaffected, there would be a correlation between kernel weight and oil content. The low oil kernels should weigh less than the high oil kernels of a segregating family. A coefficient of correlation (r) was calculated for the percent oil of the kernel and kernel weight for each exceptional family (Table 1). Families MP631, MP707, and MP784 have fairly high positive r values. There appears to be a relationship between kernel weight and oil content in these families, and the lower oil content in the segregants may be due entirely or partially to decreased germ size. Families MP663 and MP806 have very low r values and this indicates that there is no significant correlation between kernel weight and oil content. The low oil content segregants should have the same percent germ of the kernel as their high oil siblings. Thus the low oil kernels in these families are due to a decreased lipid concentration within the germ.
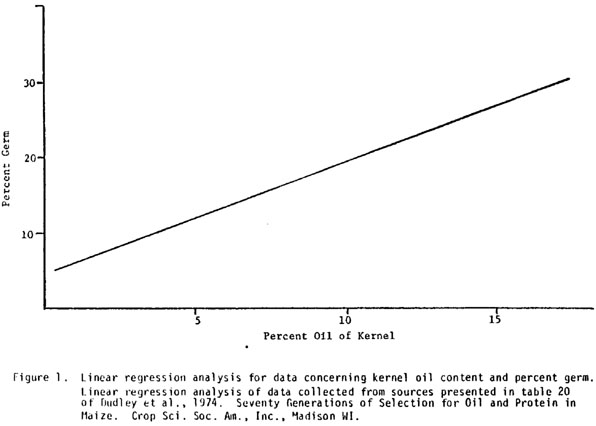
A low oil kernel (4.2%) and a high oil kernel (14.6%) were selected from family MP663 and the percent germ of each kernel was determined. The percent germ for the low and high oil kernels were 25.8% and 23.9%, respectively. I conducted a linear regression analysis (Figure 1) for percent germ and oil content from data compiled since 1898 (Dudley et al., 1974, Seventy generations of selection for oil and protein in maize, Crop Sci. Soc. Am., Inc., Madison, WI). For a kernel that has 24% germ, the oil content should be approximately 14%. This agrees well with the 14.6% oil of the high oil kernel from family MP663. The low oil segregant has 4.2% oil and if this decrease in oil content was due entirely to a decrease in germ size, then one would expect a kernel with about 10% germ (Figure 1). This last series of relationships was verified by the lack of correlation between kernel weight and oil content. Thus, these data indicate that the low oil segregants of families MP663 and MP806 are the result of a mutation that affects oil concentration in the germ and not the amount of germ in the kernel.
To determine if the low oil content characteristic (loc) would act in a Mendelian fashion after a second generation, I planted kernels from family MP663. The kernels with low oil content (X = 4.54%), the presumptive homozygous recessives, produced albino seedlings. Albinism may be pleiotropic with loc or it may be a linked independently induced mutation. The presumptive heterozygote (Loc/loc) and homozygous dominant (Loc/Loc) kernels with high oil content (X 13.40%) produced normal seedlings. Each plant was self-crossed and 50 kernels from each cob were analyzed with NMR to detect segregation of loc. Four of the seven plants were heterozygous and loc segregated in a 3:1 ratio. Data for one heterozygote, family MP4049-1, is presented in Table 1. The r for kernel weight and oil content was only 0.06. There is no correlation between these two characteristics as was the case in the parental family MP663.
I speculate that loc is expressed in homozygous recessive embryos as a severe reduction in lipid content. In the sporophyte this lipid defect affects the chloroplasts resulting in an albino seedling. This speculation predicts that a single gene regulates embryo lipids (primarily triglycerides) and sporophyte lipids (primarily polar lipids). I further speculate that loc may be a regulatory gene rather than a structural gene. Studies are in progress to map the gene and to characterize the biochemical effects of loc in the embryo and sporophyte.
Michael J. Plewa
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}