The genetics of fertility restoration for the cms-C types of cytoplasms has not been extensively studied. Recently we have converted a series of 40 sources of cytoplasmic male sterility into approximately 30 inbred lines and have studied the degree of male sterility (Gracen and Grogan, Agron. J. 66:654-657). Sixteen of the inbreds formed relatively stable male sterile combinations with either the cms-T or cms-C cytoplasms (Table 1). In our initial studies, cytoplasms C and RB comprised the C group. Subsequently we have collected several additional cytoplasms (El Salvador or ES, Bb, and Panama or PR) that seem to fall into the C group. Recently, we have attempted to determine:
(1) the number of restorer genes that interact with "C group" cytoplasms,
(2) whether or not the C, Bb, ES, PR and RB cytoplasms are identical with respect to their partial and full restoration patterns, (3) the genotypes of various inbred lines for the cms-C restorer genes,
(4) and the nature of any interactions that occur between "C group" restorer genes and those of "T group."
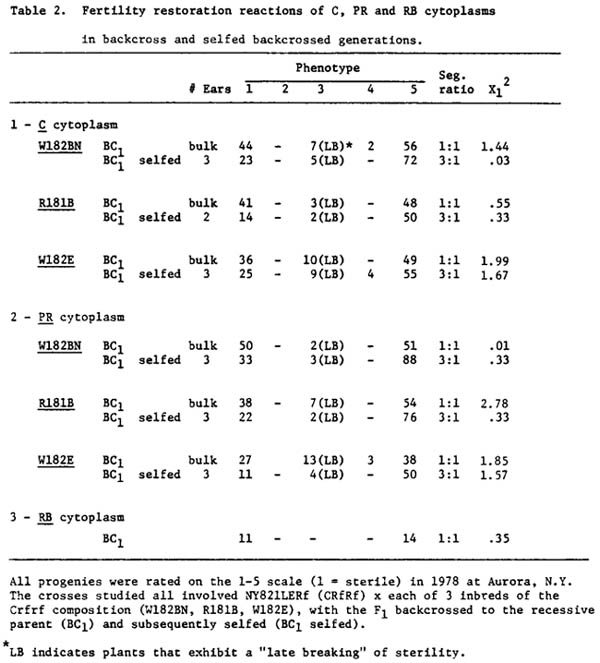
Several crosses have been made and evaluated for degree of male-sterility. Additional progenies are now under study in a Florida winter nursery. Our preliminary results show that restoration of C, RB and PR cytoplasms follows a sporophytic pattern of restoration when the line NY821LERf is used as a source of RF genes (Table 2). The data also indicate that NY821LERF is homozygous dominant for CRf factors.
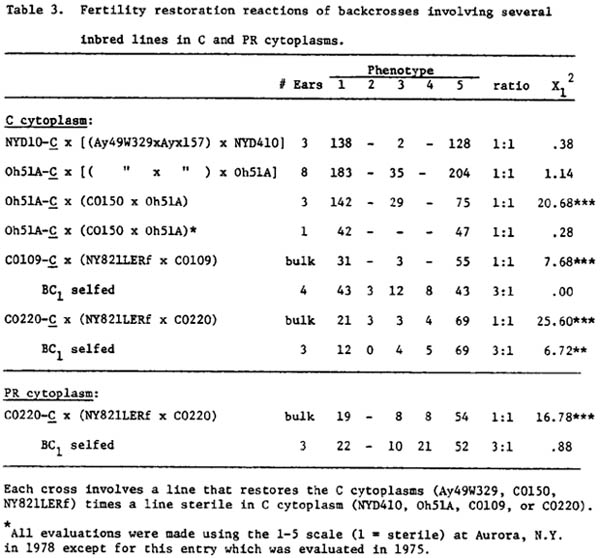
Also, the data indicate that there is apparently a single, major restorer gene for C, PR & RB cytoplasms segregating in crosses involving NY821LERf and/or Ay49W329 with recessive rf types (Tables 2 and 3). All of the BC1 progenies involving these two lines give a good fit to 1:1 ratios and the selfed BC1 progenies (with NY821LERf) give a good fit to a 3:1 ratio. In all of these crosses there are relatively few partially fertile plants and most of these are late breakers.
The segregation patterns of CO150 (as a restorer line) and CO109 and CO220 (as sterile lines) in C and PR conversions give conflicting data for a one gene segregation pattern. There may be more than one nuclear gene interacting in these crosses to condition partial or full sterility. CO150 gave a good 1:1 ratio with Oh51A-C in 1975 (ear #1 gave 42 MS:47MF). In contrast in 1978 at Aurora nursery, three ears of the same BC1 gave too many MS plants to fit a 1:1 ratio. The same difficulties are found in CO109 and CO220 crosses where progenies are characterized by too many sterile or partially fertile anthers. Since environment influences the degree of partial fertility expressed, additional studies of these apparent differences from a single gene ratio are needed. A selection program is underway to establish dependable and stable restorer CO109 and CO220 lines in a backcross program of Rf conversion.
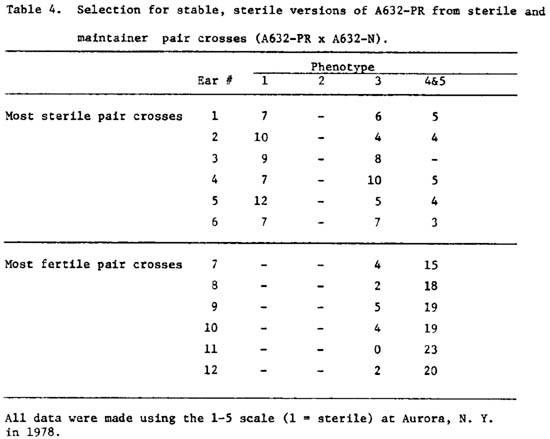
In order to facilitate use of the "C group" cytoplasms in hybrid seed production, several widely used inbred lines have been converted to these cytoplasms. Some inbred lines, such as Minn. A632, typically exhibit a partial restoration pattern in these cytoplasmic sources. All A632-C, Bb, ES, PR and RB conversions are partially restored in certain environments. PR cytoplasm appears to be the most stable source of cyto-sterility. A selection program of A632-PR conversions to try to produce a stable, male sterile line is underway. Sixteen individual plants of A632-PR genotype were crossed with individual fertile (maintainer) A632-N plants. The male parents were also selfed. In 1978 in New York, 47 of these individual A632-PR x A632 pair-cross progenies were observed. Table 4 illustrates the performance of 6 of the most sterile and 6 of the most fertile pair-cross progenies. We must point out that there are two distinct classes: (1) plants that did not exsert any anthers during the whole observation period, and (2) plants that exserted from a few anthers to nearly fully fertile tassels. One generation of selection seems to have been efficient in the selection of sterile combinations. Additional crosses were made between these sterile A632-PR plants and their sister A632-N rows last summer. Equivalent crosses were made between the most fertile A632-PR plants and their sister A632-N rows. These individual second generation progenies are under observation in a Florida nursery.
A study of environmental effects on partial fertility is also underway in greenhouse and environmentally controlled chambers. A comparative study of environmental effects of Florida and New York field environments, greenhouse and controlled growth chamber conditions will hopefully give us some concept of the role of environment vs. genetic differences for partial restorer genes in C cytoplasm restoration.
A. Kheyr-Pour, V. E. Gracen and H. L. Everett
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}