It has been suggested (M. P. Maguire, MGN 51:86-87, 1977) that maintenance of dyad integrity between anaphase I and anaphase II may depend upon the same mechanism as that which seems responsible for chiasma maintenance during the period between loss of the synaptonemal complex and anaphase I, namely sister chromatid cohesiveness. As observed cytologically at anaphase I in normal material, dyads are composed of four, usually highly condensed, chromatid arms which tend to diverge from tightly paired sister centromere regions. During interkinesis there is some decondensation of these arms followed by their recondensation during prophase II. During early prophase II in good preparations the four arms of each dyad can be traced.
If sister chromatids tend to cohere until anaphase II, except where they were separated at anaphase I as a result of crossing over, dyads would be expected to vary in their prophase II configuration in accordance with the position of their most proximal crossover in each arm. That is, a region of cohesiveness would be expected to extend from the centromere into each arm to the position of the most proximal crossover; an arm with no crossover should be cohesive throughout; a variety of complex configurations can be imagined for arms with multiple crossovers.
Camera lucida drawings were made of the dyads of 191 cells at early prophase II from four plants from diverse stocks (source indicated below). Measurements were made of the four arms of each dyad and of the regions where arms appeared to be cohesive. Where corresponding arms differed in length, the average length of the two was assigned to both. Total chromosome length was calculated for each cell; the extent of the apparently fused region at the intersection of the arms in each dyad (assumed to be the centromeric region) was divided by the total chromosome length of the cell in which it occurred. This operation was designed to give a value for extent of sister chromatid cohesiveness in this region adjusted for stage advancement, so that comparisons could be made among cells. The means and standard deviations for this value for the four plants are very similar:
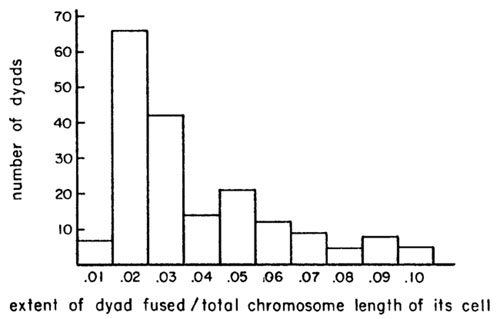
The overall mean of the extent of this fused region per dyad divided by the total chromosome length of the cell was .04 with standard deviation .0228, or an average of 20% per chromosome arm. Frequency distributions of classes of dyads with various extents of fusion/total length are shown in the following histogram.
Finally a rank difference correlation test of mean extent fusion versus total chromosome length of the cell was performed in search of a possible dependence of fusion extent on stage advancement. The rd from this calculation was -0.11 (not significant), suggesting no evidence here for such a dependence.
These results are not in accord with predictions of the hypothesis, if the genetic map is evenly distributed along the physical length of the chromosomes, and prophase II condensation is also even along chromosome length. In fact, if as usually anticipated, crossing over tends to be inhibited in centromere regions, the fit is even worse. If the hypothesis is to be seriously considered, the most reasonable speculation might be that sister chromatid cohesiveness tends to be lost to some extent proximal to crossovers during the anaphase I to prophase II interval.
Marjorie Maguire and Julia de la Cerda
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}