In a previous report (1977, MGCNL 51:13) it was noted that scutellar alcohol dehydrogenase (ADH) levels in a 1, 2, 3, 4 dosage series of 1L did not follow a direct proportionality to dosage even though the structural locus was varied. Among the possible explanations of this phenomenon, two were discussed. One interpretation is that the amount of enzyme produced is limited by an Adh specific limiting factor (see Genetics 67:411). This factor would be produced elsewhere in the genome and specifically limit the activity of the Adh genes. Thus the level of enzyme would not depend upon the number of structural genes present. A second explanation is that an inverse effect of varying 1L similar to that observed for glucose 6-phosphate, 6-phosphogluconate and isocitrate dehydrogenases would cancel a positive gene dosage effect of Adh. That is, in monosomics a reduction in ADH expression due to a reduced number of structural genes is compensated by an increased expression observed for specific enzymes. Likewise, in trisomic scutella any expected elevation in activity due to an increased number of structural genes would be cancelled by a reduction caused by a negative trisomic effect.
In order to further delimit the nature of ADH compensation, the following experiments were performed. It was reasoned that if ADH compensation were due to a cancellation of a gene dosage effect by a negative aneuploid effect, the following could be demonstrated: (1) smaller segmentally aneuploid regions that include the Adh locus should demonstrate a dosage effect for ADH; (2) a 1, 2, 3 dosage series of a portion of 1L that does not include the Adh locus should show an increase for ADH in the monosomic and a decrease in the trisomic. The magnitude of aneuploid changes must be such that when the results of 1 and 2 are multiplied together they would give the 1L observed values.
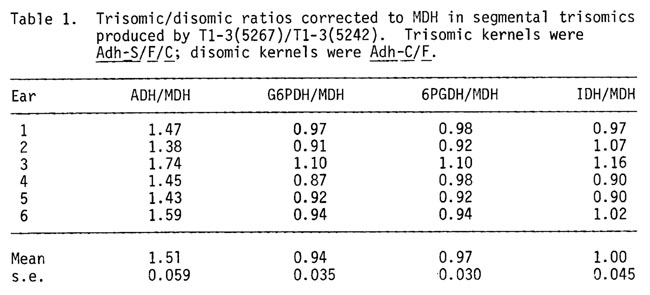
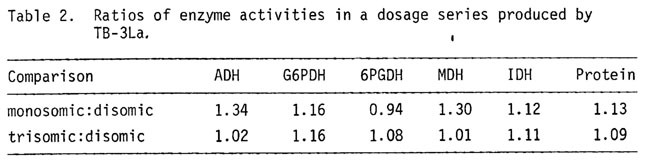
As a partial test of a dosage effect for ADH in smaller aneuploid regions, ADH, MDH (malate dehydrogenase), G6PDH (glucose-6-dehydrogenase), 6PGDH (6 phosphogluconate dehydrogenase), IDH (isocitrate dehydrogenase) and total protein were measured in trisomic scutella vs disomic scutella from kernels resulting from a cross of a female, heterozygous for T1-3(5267) and T1-3(5242), by a normal male. This cross is described in greater detail in the previous report. Scutella of Adh-S/C/F genotype (trisomic) were compared to Adh-C/F (disomic) for their ADH/MDH ratios. These data are given in Table 1. The mean (+ s.e.) 3 dose/2 dose ratio for ADH is 1.51 + 0.059 and is clearly proportional to gene dosage in this segmental trisomic which is considerably smaller than the trisomics produced by TB-1La. The values observed here differ from the 1.16 mean trisomic/disomic ratio found in whole arm comparisons (Birchler, Ph.D. thesis, Indiana). The straightforward interpretation is that Adh shows a gene dosage effect in this trisomic-disomic comparison. Other possibilities include the following. The small partial 3L trisomic region has a positive effect on ADH levels. Since ADH levels (Table 2) in a 3L(TB-3La) dosage series do not show a trisomic effect, this seems unlikely. Secondly, a gene for an Adh limiting factor is encoded in the 1L or 3L region that is trisomic. The increase in ADH would then be due to an increase in its limiting factor. This possibility would require that an inverse effect would work on the limiting factor locus or its product in the aneuploids produced by TB-1La in order to produce the ADH compensation observed.
Perhaps segmental trisomics involving other 1L breakpoints as well as other chromosome arms than 3L as the concomitant trisomic region will test these possibilities. Translocation combinations involving the region of 1L that will triplicate Adh as well as regions of 8L and 10L have been made and will be tested.
Another experiment designed to test whether a dosage effect for ADH is demonstrable in otherwise monosomic, disomic and trisomic scutella involved crossing the T1-3(5267)/T1-3(5242) heterozygote by TB-1La. This cross will add an extra dose of Adh to scutella monosomic, disomic and trisomic for the region of 1L involved in TB-1La.
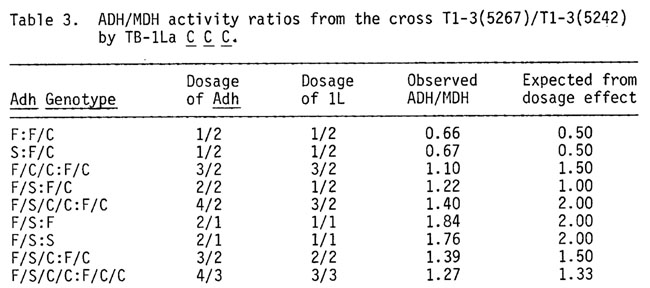
In the interchange heterozygote mentioned above, Adh is also heterozygous for F and S alleles. Thus when it is used as female three types of viable gametes, with respect to Adh genotype, are formed: F, S and F/S. The F/S gametes are duplicate for the "between the breakpoints" regions of 1L and 3L. This 1L region includes the Adh locus. Thus if the interchange heterozygote is crossed by a plant hyperploid (1 B1 B1 1B) for TB-1La and homozygous for Adh-C, which will produce gametes with 0, 1 or 2 doses of 1L marked by this different Adh variant, the resulting zygotes are as follows: F/-, S/-, F/S/-, F/C, S/C, F/S/C, F/C/C, S/C/C and F/S/C/C. The first three classes of zygotes are monosomic for 1L; the second group of three have two doses of the region involved in TB-1La; the last three are trisomic for 1L. In each group the one with F and S have an extra dose of 1L 0.72-0.90. Thus one can compare monosomics to monosomics plus an extra Adh; disomics to disomics plus an extra Adh; and trisomics to trisomics with an extra Adh as well as the usual monosomic, disomic, trisomic comparison--all from the same ear. All classes of zygotes can be distinguished from the others on the basis of which Adh alleles are present and their isozyme band ratios. ADH was measured in the following classes of scutella: F/-, S/-, F/S/-, F/C, F/S/C, F/C/C and F/S/C/C. The ratios of ADH activities in various comparisons are given in Table 3.
The ADH expression in monosomics shows a typical level compared to the disomic (0.66). Also in the whole arm trisomic, ADH is compensated, showing 1.10 as the 3/2 dose ratio. The two doses of Adh in the monosomic class show close to twice (1.80x) as much ADH as found in the one dose monosomic. The three doses of Adh in the disomic show 1.39 times as much ADH activity as present in the disomic. Thus from kernels on the same ear the small trisomic region around Adh shows a significantly greater ADH expression than in the larger trisomic region (small/ large = 1.39/1.10). Furthermore, if an extra Adh dose is added to the trisomic, a dosage effect is observed. One would expect the 4/3 dosage effect to be 1.33. The observed value was 1.27. If one compares these four doses in a whole arm trisomic to two doses in the disomic, one finds a ratio of 1.40. For a strict dosage effect, a ratio of 2.00 would be expected. Thus the trisomic condition must reduce the total output of the four doses to 70% of the expected value from a dosage effect.
From these data it appears that the expression of ADH is positively affected by the dosage of the Adh locus and negatively correlated with the dosage of 1L. At any level of aneuploidy, an increase in Adh dosage shows a dosage effect reasonably close to the expected effect. However, if the dosage of 1L is varied, there is a tendency for ADH compensation.
Further work on this problem will involve testing compound TB-A's which have replaced the terminal end of 1L with some other phenotypically marked region as well as placing the terminal end of 1L (using Adh as a genetic marker) onto other TB-A's. The compounds constructed will test several lengths of 1L proximal to Adh in a 1, 2, 3 dosage series as well as distal regions including the Adh locus. This will leave only the region distal to Adh to be tested independently of Adh dosage.
In conclusion, this progress report has noted evidence that suggests, but does not prove, that the compensation of ADH in the dosage series produced by TB-1La is due to a cancellation of a gene dosage effect by an inverse effect produced by 1L. Further studies as noted above will be conducted in order to discriminate between the other mentioned possibilities.
James A. Birchler
Return to the MNL 52 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}