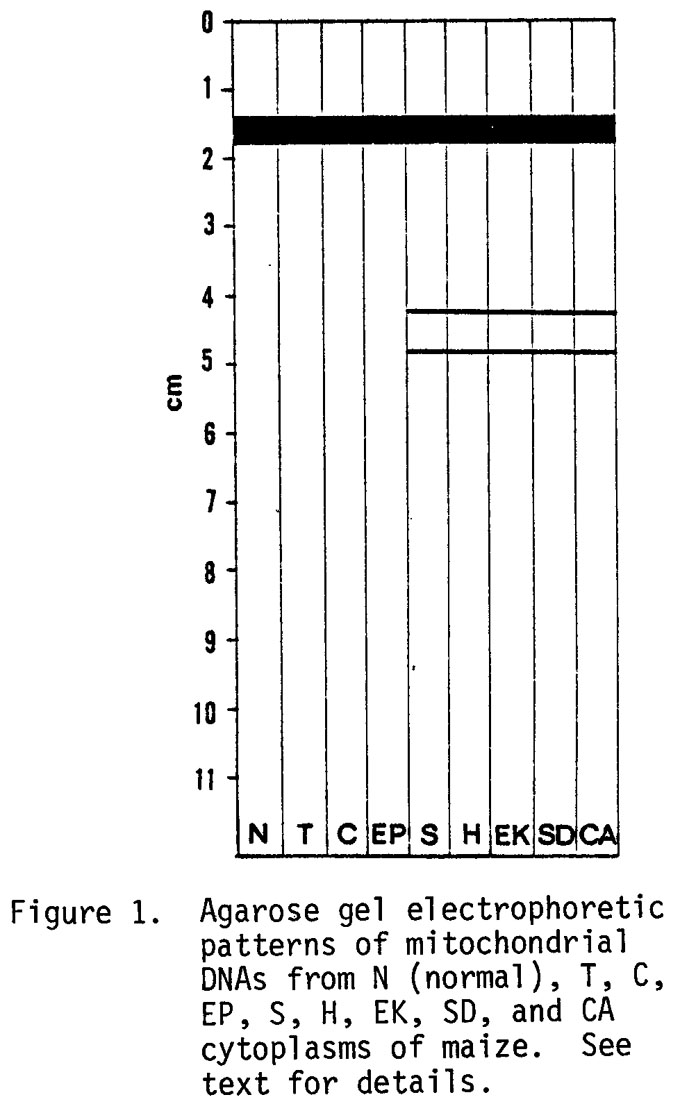
When maize mitochondrial DNA (mtDNA) was fractionated by agarose gel electrophoresis, it showed a single broad band of high molecular weight DNA at about 1.5 cm (Figure 1) for all cytoplasms studied (N, T, C, EP, S, H, EK, SD, and CA). Interestingly, members of the S group (S, H, EK, SD, and CA) manifested two additional bands which we have designated S-S (slow) and S-F (fast) DNAs. These unique bands and all bands described herein were abolished by deoxyribonuclease and unaffected by ribonuclease. The mtDNA was prepared by CsCl-ethidium bromide preparative ultracentrifugation as previously described (Science 193:158-160). The S-S and S-F DNAs have been obtained from upper and lower bands of CsCl-dye gradients which indicate that the DNA exists as open-circular or linear and supercoiled molecules. The two unique DNA bands associated with the S group of cytoplasmic male steriles (cms) are the subject of this report.
The S cms was studied in the nuclear backgrounds WF9, Mo17, FR37, WF9 x W64A, N6 x W64A, and FR37 x K21; EK, H, and SD cms were in Mo17; and CA cms was in W64A. All the above types yielded the two unique DNAs associated with the S group of cms. The cross FR37 cms S x K21 was restored for pollen fertility, but nonetheless contained the unique DNAs S-S and S-F. To further establish the asso- ciation of the two unique DNAs with the S cytoplasm, we looked for the unique DNAs in 10 N (normal) cytoplasms, F6, F44, T204, W64A, WF9, 61M, A619 x A632, B37 x NC236, NC7 x T204, and B37 x NC236; in 5 sources of T cytoplasm, F44, T204, W64A, B37 x NC236, and NC 7 x T204; in 3 sources of C, Mo17, WF9, and B37 x NC236; and in 1 source of EP, A619 x A632. The unique DNAs present in the S group of cms were not found in the N, T, or C cytoplasms.
Molecular weights of the S-S and S-F DNAs were estimated by two techniques, gel electrophoresis and electron microscopy (Table 1). Molecular weight determinations were made by electrophoresis in 1.5% agarose gels, with Eco RI-digested lambda and adenovirus type 2 DNA as molecular weight markers. Since the molecular weight markers were linear molecules, Hind III digests of S-S DNA were electrophoresed to better estimate the size of S-S DNA. Three fragments were obtained, with molecular weights of 1.47, 1.22, and 1.02 x 106 daltons, for a total molecular weight of 3.71 x 106 daltons. Hind III digestion of S-F DNA produced two fragments with molecular weights of 2.30 and 1.12 x 106 daltons, resulting in a total molecular weight estimate of 3.42 x 106 daltons. No apparent variation in molecular weight of the S-S and S-F DNAs was detected among the S, CA, EK, SD and H cytoplasms.
The two unique DNAs associated with the mtDNA from the S cytoplasm were studied by electron microscopy. These DNAs were isolated by electrophoretic separation of the upper DNA band of a CsCl-ethidium bromide gradient, because our yield of lower band (supercoiled) DNA was insufficient for this study. Most of the DNA molecules isolated in this manner and examined by electron microscopy were linear except for a few circular molecules. Although the upper band from a dye-CsCl gradient contains circular as well as linear molecules, we have experienced consistent difficulty in obtaining high yields of circular molecules after gel electrophoresis.
The size of the linear molecules of S-S DNA was determined by measuring their length relative to the length of open-circular fX174 R II DNA which was used as an internal standard in the same spreading. The ratio of S-S DNA length to fXV174 was 1.13. The molecular weight of S-S DNA is 3.84 x 106 when a molecular weight of 3.4 x 106 is used for fX174 DNA. Similarly, the size of S-F DNA was obtained by measuring length relative to the length of 4.67 x 106 molecular weight linear fragments from Eco RI-digestion of the bacteriophage lambda. The ratio of S-F DNA to the lambda DNA fragment was .73 so that the molecular weight of the S-F DNA is 3.41 x 106. These molecular weights of 3.41 and 3.84 x 106 for S-F and S-S respectively are comparable with those determined by gel electrophoresis of 3.42 and 3.71 x 106 for S-F and S-S, respectively.
The two sizes of low molecular weight, circular DNAs from the S cytoplasm seem to be distinct species, as demonstrated by electrophoresis of open circular and linear molecules, and by electron microscopy. Mitochondria from N and T cytoplasms also contain low molecular weight circular DNAs (Shah, et al. in preparation) but electrophoresis does not elaborate these as distinct bands. Apparently, in the N and T cytoplasms, small circular DNAs do not occur with sufficient frequency for electrophoretic visualization. The mtDNA of maize, whether from N, T, or S cytoplasm, is heterogeneous in contour length; it ranges from mini-circles (less than 3 µm) to circles as large as 33 µm. The reason for this intermolecular heterogeneity, and its relation to the unique S DNAs, is not clear. Perhaps the latter DNAs are a special form of intermolecular heterogeneity.
A most interesting characteristic of the S cytoplasms is their instability which has been described in detail by Laughnan and his associates (Theor. Appl. Genetics 43:109-116; Genetics 71:607-620). It is tempting to speculate that the circular DNAs described here are in some way related to the unstable nature of the S male-sterile cytoplasm, and in fact, may represent elements described by Laughnan and Gabay (In Birky, C. W., Jr., Perlman, P. S. and Byers, T. J., eds., Genetics and Biogenesis of Mitochondria and Chloroplasts, Ohio State University Press, pp. 330-349). They hypothesized an episomal element to explain the occurrence of fertility restoration by a nuclear gene, with the corresponding requirement that the cytoplasmic element be capable of transposition to, and stabilization in, a chromosomal site. Our data do not permit a distinction among an episomal, viral or other extraneous agent in our S cytoplasm stocks.
To date our evidence suggests that the unique DNAs associated with the S cytoplasm are confined to the mitochondria. We have interpreted our data as circumstantial evidence of a mitochondria-sterility association, but a causal relationship between the unique mtDNAs of the S cytoplasm described in this paper and the expression of male sterility in maize remains to be established.
C. S. Levings III, D. R. Pring,* W. W. L. Hu and D. H. Timothy
*ARS, USDA and Department of Plant Pathology, University of Florida
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}