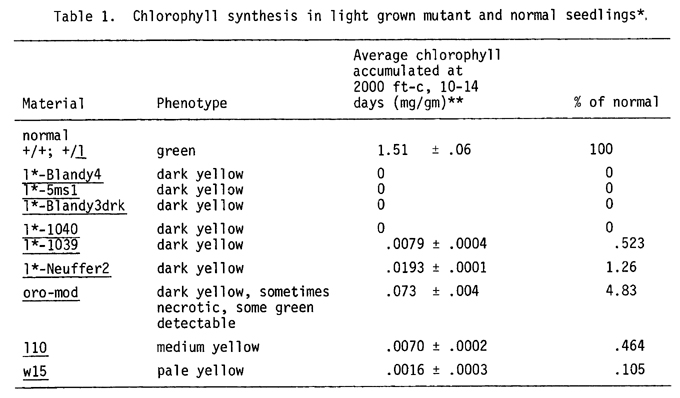
The porphyrin molecule is ubiquitous in nature and its synthesis has been studied in a variety of organisms. The study of mutants has been instrumental in explicating porphyrin and specifically chlorophyll biosynthesis. In higher plants chlorophyll synthetic mutants have been studied extensively only in barley (see von Wettstein et al., 1974, Science 184:800 for information and references). The techniques used in studying barley mutants are directly applicable to corn. A preliminary report of some luteus mutants in corn is presented here. Figure 1 presents a summary of the chlorophyll pathway showing possible regulatory molecules and mutants of corn and barley affecting various steps in the pathway. Table 1 presents the phenotypes of the mutants discussed. Only orobanche-modified (oro-mod) develops any visibly detectable chlorophyll at 2000 ft-c of light, albeit several do accumulate a small amount of chlorophyll not visible to the naked eye.
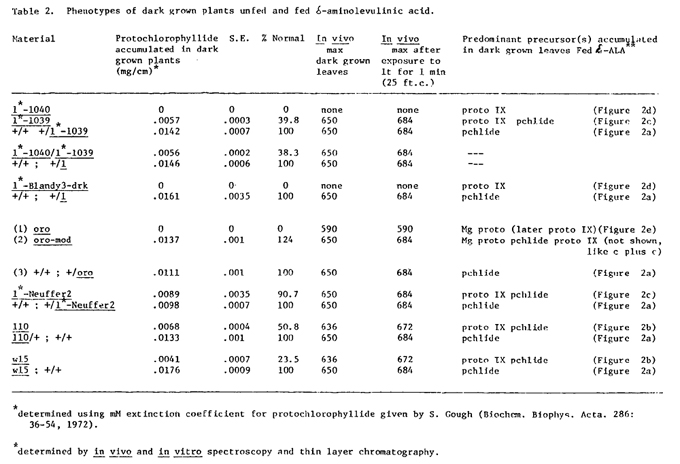
Dark grown material is particularly useful in determining the defective step in chlorophyll synthesis since higher plants are naturally blocked in the conversion of protochlorophyllide to chlorophyllide. The pathway is further inhibited by a feedback mechanism preventing d-aminolevulinic acid synthesis. In normal plants this results in accumulation of a small amount of protochlorophyllide which is attached to holochrome protein (Rebeitz, C. and Castelfranco, P. Ann. Rev. Plant Physiol. 24:129-172, 1973). In vivo at least two types of protochlorophyllide holochrome are present. One, absorbing maximally at approximately 650 nm, is immediately convertible to chlorophyllide on exposure to light. The other is non-photo-transformable and has an absorption maximum of 636 nm. A variety of dark grown mutants has been examined by in vivo and in vitro spectroscopy (Table 2). The mutants discussed here can be divided into three physiological categories although genetically these classes overlap: 1) completely blocked mutants which accumulate no protochlorophyllide, 2) leaky mutants which accumulate protochlorophyllide 650, and 3) leaky mutants which accumulate reduced protochlorophyllide absorbing maximally at 636 nm.
The completely blocked mutants include l*-Blandy4 and alleles (l*-EMS1 and l*-Blandy3drk); l*-1040 ; and some recently found stocks of oro. The former two produce no detectable chlorophyll precursors while the latter produces a small amount of Mg-protoporphyrin probably corresponding to the amount of protochlorophyllide produced in normal leaves. We interpret this to indicate that protoporphyrin IX is involved in regulating the porphyrin pathway. The small amount of either Mg-protoporphyrin in oro or protochlorophyllide in normal seedlings probably results from competition between synthetic enzymes and regulatory molecules for the small amount of protoporphyrin IX produced. Presumably the heme pathway also competes for the same protoporphyrin IX, however, this has not been clearly established in higher plants.
The leaky mutants which accumulate protochlorophyllide 650 include oro-mod, l*-Neuffer2 and l*-1039, a leaky allele of l*-1040. Orobanche was originally found in a leaky background and was placed in this category. However, this leakiness may be due to the presence of a modifier gene. The recent observation of 3 non-leaky plants out of 12 mutant seedlings tested from a self pollinated ear approximates a 3:1 ratio among mutants, expected if the non-leaky oro plants are homozygous for a recessive allele at an independent modifier locus. Further genetic tests are necessary to determine the nature of the modifier but it seems reasonable to believe that basically oro is a non-leaky mutant. Both l*-Neuffer2 and l*-1039 accumulate protochlorophyllide 650 which is photoconvertible to chlorophyllide 684 on exposure to light: the former accumulates an approximately normal amount while the latter accumulates about 40% of the normal level of protochlorophyllide. It is interesting to note that the heterozygote l*-1040/l*-1039 has a leaky phenotype producing the same amount of protochlorophyllide as the homozygous leaky mutant. These mutants probably represent one of the following: 1) leaky enzymes of chlorophyll synthesis, 2) mutants in structural elements to which precursors and/or enzymes must bind or 3) mutants whose expression is partially suppressed by modifier genes. In any case a small amount of protochlorophyllide can be synthesized when the demand is low, as in darkness, but under normal lighting conditions they cannot meet the demand for chlorophyll.
The third class of mutants produces reduced protochlorophyllide which absorbs maximally at 636 nm and is thus not photoconvertible to chlorophyllide 684. There are many mutants with this phenotype, including l10 and w15, which may result from defects in any of the undefined processes necessary for normal chloroplast development.
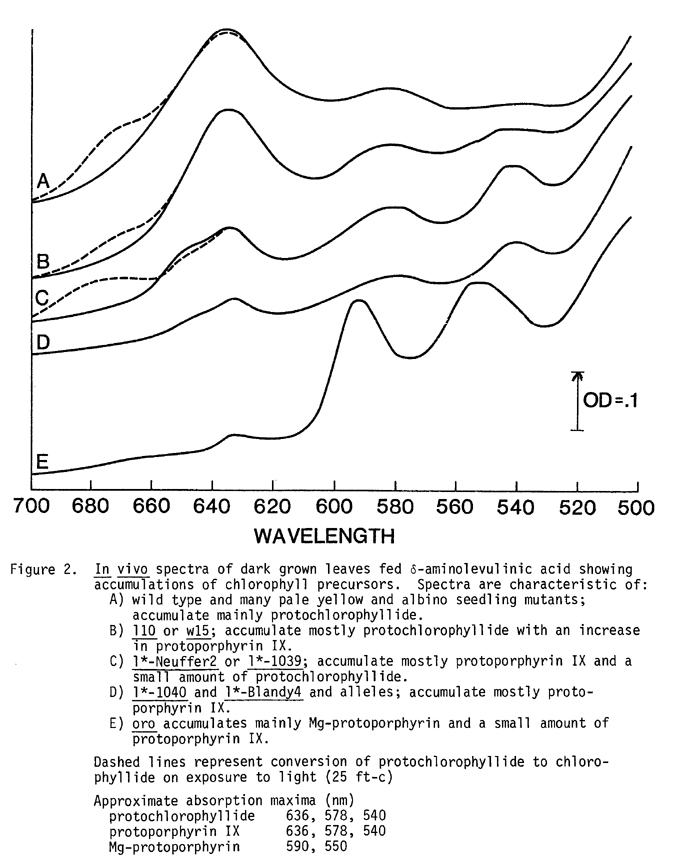
Although protochlorophyllide synthesis is generally tightly regulated the regulatory step can be bypassed by feeding detached shoots d-aminolevulinic acid (d-ALA) (.01M in distilled H2O, 24 hrs). After this treatment, normal plants accumulate a large amount of protochlorophyllide 636 as well as a small amount of protoporphyrin IX (Figure 2, A). Examination of mutants after feeding d-ALA by in vivo spectroscopy and thin layer chromatography makes possible identification of precursors built up between d-ALA and protochlorophyllide.
The results are summarized in Figure 2 and Table 2. There are many pale yellow and albino mutants that accumulate a normal level of protochlorophyllide when fed d-ALA (Figure 2, A). Several mutants, including l10 and w15, accumulate a small amount of protoporphyrin IX as well as protochlorophyllide (Figure 2, B). These lesions are not believed to be directly involved in chlorophyll synthesis but result in the inability of chloroplasts to stabilize chlorophyll. The leaky mutants l*-Neuffer2 and l*-1039 accumulate mostly protoporphyrin IX, but some protochlorophyllide is accumulated (Figure 2, C). Completely blocked mutants l*-Blandy4 and alleles, and l*-1040, accumulate predominantly protoporphyrin IX when fed d-ALA (Figure 2, D). No protochlorophyllide has been detected in these seedlings even after feeding. Figure 2, E, which represents oro fed d-ALA for 7 hours, demonstrates accumulation of predominantly Mg-protoporphyrin. After 24 hours a large amount of protoporphyrin IX was also accumulated. Leaky oro (oro-mod) seedlings accumulate Mg-protoporphyrin as well as protoporphyrin IX and protochlorophyllide.
In summary: Several mutants specifically involved in chlorophyll synthesis either directly as chlorophyll synthetic enzymes or indirectly as structural chloroplast components have been analyzed. Three loci are defective in conversion of protoporphyrin IX to Mg-protoporphyrin: 1) l*-Blandy4 and alleles; 2) l*-1040 and allele l*-1039; and 3) l*-Neuffer2. The former two can be completely blocked but no non-leaky allele of l*-Neuffer2 has been found. One mutant, oro, is blocked in the conversion of Mg protoporphyrin to protochlorophyllide. Several other mutants, including l10 and w15, are probably defective in chloroplast synthesis. No blocks prior to protoporphyrin IX such as xan-u in barley have been found. However, the number of mutants examined in corn is relatively small compared to the barley study. The discovery of a putative modifier gene in oro which allows synthesis of protochlorophyllide is novel and will be examined further. It is possible that other leaky mutants are the result of similar modification.
Peter Mascia and D. S. Robertson
Return to the MNL 51 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}